Contribution à l'étude du zooplancton du lac Kivu : cas du bassin est au large de Kibuye( Télécharger le fichier original )par Theoneste NZAYISENGA Université Nationale du Rwanda - Bachelor of Science in Biology 2007 |

IV.1.2.2 L'oxygène dissousLes profils verticaux d'oxygène dissout durant toute la période de notre échantillonnage sont présentés à l'annexe 2. Au cours de notre travail l'oxygène dissous dans l'eau de surface a varié entre 10mg/l et 6mg/l. 116. 5 LL Comme la température, l'oxygène dissous diminue progressivement avec la profondeur - =-1 dans la colonne d'eau suite á l'absence des mouvements de brassage de l'eau. Ces mouvements sont influencés par les vents qui soufflent sur le lac. IV.1.2.3 La conductivité Les profils verticaux de la conductivité de l'eau du lac Kivu durant toute la période de notre échantillonnage sont présentés à l'annexe 4. Contrairement aux autres paramètres, la conductivité ne montre pas d'importantes variations. Dans l'eau de surface, les valeurs de la conductivité mesurée étaient comprises entre 1000 et 1500 us/cm. Cependant, la conductivité croît dans les couches les plus profondes de l'eau du lac Kivu (à partir de 60 m) suite à des concentrations importantes en sels dissous. IV.1.2.4 Le pH Les profils verticaux du pH dans la colonne d'eau du lac Kivu sont présentés à l'annexe 3. L'observation de la figure en annexe montre que le pH de l'eau du lac Kivu ne varie pas beaucoup. Les valeurs de pH mesuré dans l'eau de surface étaient de 8 unités de pH et diminue progressivement pour devenir légèrement acide (6,6 unités de pH) à 100m de profondeur suite à l'augmentation de la concentration en ions H+ d'après la réaction suivante : CO2 + H2O ? H2CO3 ? H+ + HCO3 - - Si la concentration en CO2 est élevée ; l'équilibre sera déplacé vers la droite : H+ augmente, donc le pH diminue. - Si la concentration en CO2 est faible ; l'équilibre sera déplacé vers la gauche : H+ diminue, donc le pH augmente et HCO3 - (bicarbonates) joue le rôle de réserve de CO2. IV.1.2.5 Profondeur de la zone euphotique L'absorption du rayonnement solaire est rapide dans l'eau. La profondeur pour la quelle l'intensité de la lumière incidente est réduite à 1% de sa valeur à la surface (zone euphotique) varie de 17.4 à 33,7 m suivant les cas. La figure XIII montre les différentes valeurs prises par ce facteur durant la période de notre étude. Au lac Kivu, la limite de la zone euphotique se trouve en moyenne à 22.6m ; elle présente un maximum au mois d'aoüt (33.7 m) et un minimum en octobre (17.4 m) après le passage des vents forts qui ont occasionné le brassage des eaux du lac. Suite à de fortes vagues, la transparence n'a pas été mesurée lors de certaines sorties d'échantillonnage ; il s'agit du 12/04/05, 05/07/05, 02/08/05 et du 05/10/05.
Figure XIII : Variation de la limite de la zone euphotique dans le lac Kivu bassin de Kibuye. IV.1.3 COMPOSITION ET TAILLE DU ZOOPLANCTON IV.1.3.1 Composition du Zooplancton Le zooplancton pélagique du lac Kivu, bassin Est au large de Kibuye, est Dt 17:3 principalement composé de trois espèces de Copépodes, Tropocyclops confinis - (KIEFER, 1930), Thermocyclops consimilis (KIEFER, 1934) et Mesocyclops aequatorialis (KIEFER, 1929) ; de trois espèces de Cladocères, Diaphanosoma excisum (SARS, 1886) Alona rectangula (SARS, 1862) et Moina micrura (KURZ, 1874) ainsi que des Rotifères comprenant, Keratalla spp. et Brachionus quadridentatus (HERMANN, 1783). IV.1.3.2 Taille du Zooplancton Les échantillons du zooplancton mesuré montrent que la taille moyenne du zooplancton du lac Kivu bassin de Kibuye est de 0,4 mm. La plus grande moyenne se rencontre chez les copépodes où la taille moyenne est de 0,59 mm et leur nauplii présentent la plus petite moyenne 0,16.
Dates 12-Apr-05 1.01 26-Apr-05 0.49 10-May-05 1.37 24-May-05 0.71 7-Jun-05 0.92 21-Jun-05 0.54 5-Jul-05 1.24 19-Jul-05 0.44 2-Aug-05 0.42 16-Aug-05 0.33 13-Sep-05 1.44 5-Oct-05 3.80 25-Oct-05 0.64 Biomasse totale du zooplancton (gMSm-2) IV.1.4 VARIATION TEMPORELLE DU ZOOPLANCTON La densité totale du zooplancton (tous les taxons confondus), de même que la biomasse totale, varie considérablement. L'examen de la figure XIV et du tableau VII montre que le zooplancton pélagique du lac Kivu bassin de Kibuye a atteint son maximum le 4 octobre 2005, avec une densité totale estimée à 500*104 Ind./m2 et une biomasse estimée à 3.80gMs/m2. La plus faible densité s'observe le 16 aofit où elle est estimée à 44*104 Ind./m2. La plus faible biomasse est observée le même jour et elle est estimée à 0.33gMs/m2. Tableau V : Biomasse totale du zooplancton pélagique du lac Kivu bassin de Kibuye liSipillq ITENTEMEEE5)
Figure XIV : Variation de la densité totale du zooplancton pélagique du lac Kivu bassin de Kibuye (avril - octobre 2005) IV.1.5. ABONDANCE RELATIVE DE DIFFERENTS GROUPES DE ZOOPLANCTON P.% 7"4 (17 $ L'abondance relative des différents groupes du zooplancton du lac Kivu durant toute la période de notre échantillonnage régulier sur toute la colonne d'eau (60 m) est reprise dans la figure XV. Les copépodes dominent largement les autres groupes, avec un pourcentage d'abondance qui varie entre 84% et 92%, les cladocères viennent en 2eme position avec les pourcentages qui varient entre 7% et 16% , les rotifères était très peu représentés (moins de 2%). La faible représentation des rotifères pourrait être expliquer par le fait que la maille du filet à plancton utilisé était trop large par rapport à la taille des rotifères (100um).
Figure XV 1 9 DUDAioQ 01 O'DEoQ0DQE1111ODABH0IK ILDQ0s OLIXSIs 0X IIRSODQcAoQ pélagique du lac Kivu bassin de Kibuye V.1.6 DISTRIBUTION VERTICALE DU ZOOPLANCTON L'observation de la figure XVI, montre que la densité du zooplancton du lac Kivu varie considérablement dans la colonne d'eau et que le zooplancton est plus concentré dans la couche de 0 à 20m de profondeur. Le pic zooplanctonique suit immédiatement le pic du phytoplancton et les deux pics apparaissent lorsque la zone de mélange a été la plus grande (c'est-à-dire atteignant une grande profondeur); après le brassage des eaux du lac Kivu. Tout au long des mois d'avril, mai et début de juin, la densité du zooplancton était estimée entre 80.104 et 180.104 individus par m2. C'est au cours de la période des mois de juillet et août que la densité du zooplancton a été la plus faible (entre 50.104 et 60.104 indim2). Durant les mois de septembre et octobre, la densité du zooplancton a largement augmenté, variant de 70.104 indim2 à 500.104 indim2. La densité du zooplancton a atteint son minimum en période de saison sèche (mois d'aoüt), période pendant laquelle le zooplancton atteignait son maximum de densité normalement. Ceci pourrait être expliqué par les changements climatiques qu'a connu la sous région. Par exemple les données météorologiques collectées à la station de Gisenyi montrent que c'est au mois d'aoüt qu'on a enregistré plus de pluies (fig.IX). Signalons qu'au début du mois d'octobre, la couche allant de 40 à 60 m de profondeur qui était d'habitude la moins peuplée par le zooplancton (KANINGINI, 2003) s'est retrouvée avec plus de 100.104 indim2 en date du 4 octobre 2005.
Figure XVII : Relation entre la transparence de l'eau du lac Kivu et la moyenne mensuelle de la densité du zooplancton L'examen de la figure XVII révèle qu'en moyenne, aux valeurs les plus élevées de la de leau du lac densité zooplanctonique correspondent les valeurs les plus basses de la transparence de 0 12 l'eau du lac Kivu et inversement ; la valeur la plus élevée de la densité zooplanctonique 0 8 D tot 0 s'observe au mois d'octobre où la transparence se trouve à son minimum. Le coefficient V Transparence 4 de corrélation négatif (R= -0,71) indique une relation négative entre les deux variables. 0 Cependant, seul le zooplancton ne peut pas justifier la transparence de l'eau du lac, si A M J J A S O non la transparence trouvée au mois de septembre et
octobre ne serait pas au même
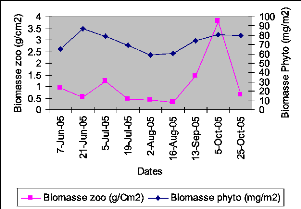
tion des se (/C co 5' 3 0. CD CP 0 0 CD4 a. zooplancton augmente sa densité lorsqu'il y a plus de nourriture (le phytoplancton majoritairement) disponible pour lui. La figure XVIII reprend une comparaison entre la variation de la biomasse zooplanctonique et celle phytoplanctonique durant une partie de notre travail sur le train.
Figure XVIII : Variation de la biomasse du zooplancton et du phytoplancton pélagique au lac Kivu bassin de Kibuye de 0 à 100m de profondeur (juin-octobre 2005) V.1.7 MIGRATIONS VERTICALES DU ZOOPLANCTON Les migrations verticales du zooplancton ont été
analysées à la fin du mois d'octobre et 90 l'autre durant l'obscurité totale, le 11 novembre 2005. La migration de chaque taxon a 70 été suivie et les copépodes adultes et leurs nauplii ont été suivis séparément. 50 v a. Cycle de 24 heures du 25 octobre 2005 (pleine lune) 0 Figure XIX : Migration verticale des copépodes du lac Kivu basin de Kibuye en pleine lune (le 25/10/2005) La fig. XIX montre que les copépodes adultes, toujours présents en grande quantité dans la couche superficielle, augmentent leur densité dans la couche de 0 à 10 m et le pic de migration verticale de ces copépodes adultes se manifeste à 18H en période de pleine lune.
Figure XX : Migration verticale des Nauplii du lac Kivu basin de Kibuye en pleine lune (le 25/10/2005) L'observation de la fig. XX montre que la migration verticale des nauplii n'est pas orientée vers la surface comme le faisaient les adultes, au contraire les nauplii migrent en profondeur est présentent le pic de leur migration à 6H du matin à une profondeur de 30m.
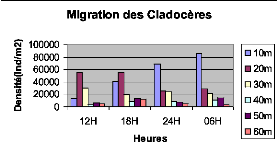
Figure XXI : Migration verticale des Cladocères du lac Kivu basin de Kibuye en pleine 40m lune (le 25/10/2005) 12H 18H L'observation de la fig. XXI montre que les Cladocères migrent vers la surface le matin à 6H et présentent le pic de migration dans la couche de 10m de profondeur.
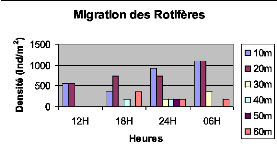
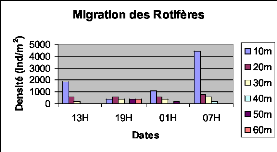
Figure XXII : Migration verticale des Rotifères du lac Kivu basin de Kibuye en pleine lune (le 25/10/2005) L'observation de la fig. XXII montre que les Rotifères migrent vers la surface le matin à 6H et présentent le pic de migration dans la couche de 0 à 20m de profondeur. b. Cycle de 24H du 08 novembre 2005 (nuit sans lune) 20m
ion des 8H 24H 06 50m Figure XXIII : Migration verticale des Copépodes adultes du lac Kivu bassin de Kibuye 40m en période de nuit sans lune (le 08/11/205) 50m 0 L'examen de cette fig. XXIII montrent que les Copépodes adultes migrent vers la surface et manifestent leur pic de migration dans la couche de 0 à 10m de profondeur à 19H en période de nuit sans lune.
Figure XXIV : Migration verticale des Nauplii du lac Kivu bassin de Kibuye en période de nuit sans lune (le 08/11/2005) La fig. XXIV montrent que les nauplii migrent en profondeur et présentent le pic dans la couche de 20 à 30m de profondeur pendant la nuit 19H et 01H en période de nuit sans lune.
Figure XXV : Migration verticale des Cladocères du lac Kivu bassin de Kibuye en 30m période de nuit sans lune (le 08/11/2005) 40m L'examen de la fig. XXV montre que les Cladocères restent présentes en grande quantité dans 60m Dates la couche de 0 à 10m. Les mouvements migratoires des Cladocères en période de nuit sans lune ne sont pas remarquables puisqu'ils sont stimulés par la lumière. Donc la migration des Cladocères du lac Kivu bassin de Kibuye varie suivant la position de la lune. C'est-à-dire qu'en pleine lune ils migrent et au crépuscule ils ne migrent pas.
Figure XXVI : Migration verticale des Rotifères du lac Kivu bassin de Kibuye en période de nuit sans lune (le 08/11/2005) L'observation de la fig. XXVI, montre qu'en période de nuit sans lune les Rotifères migrent vers la surface le matin à 7H. D'une façon générale la migration du
zooplancton du lac Kivu bassin de Kibuye (à 10m densité pour les copépodes se manifeste à 18H en pleine lune et à 19H en période de 20m nuit sans lune. Nous pouvons conclure que les copépodes du lac Kivu bassin de Kibuye 4 migreraient vers la surface pendant la nuit. Quant au nauplii, ils se concentreraient en 50m 13 profondeur à 30m surtout pendant la nuit. Les cladocères eux migreraient vers la surface 60m Dates le matin en pleine lune mais en période de nuit sans lune ils semblent ne pas migrer de façon significative. Les rotifères eux migreraient vers la surface le matin sans toutefois être influencés par la position de la lune. IV. 2 DISCUSSION ET INTERPRETATION Les échantillons du zooplancton et des paramètres physico-chimiques de l'eau du lac Kivu bassin de Kibuye ont été collectés d'avril à novembre 2005. La transparence moyenne de l'eau du lac Kivu bassin de Kibuye était estimée à 6,8m. La limite de la zone euphotique variait entre 17 et 34m de profondeur. Cette zone est plus large au mois de juillet et août, tandis qu'en septembre et octobre elle est la plus petite. La grandeur de la zone euphotique varie avec la concentration des matières en suspension tel que le plancton. La remontée des nutriments est due aux vents forts qui provoquent les mouvements de brassage dans la colonne d'eau, et occasionne un bloom planctonique. Ce qui provoque une diminution de la transparence de l'eau. Au bassin de Bukavu, KANINGINI et al., 2003 ont trouvé une transparence inférieure à celle trouvée à Kibuye parce que au bassin de Bukavu les eaux du lac se mélangent complètement jusqu'à une profondeur de 100 m tandis qu'à Kibuye la zone de mélange ne dépasse pas 70m de profondeur. C'est la morphologie du bassin versant qui est à l'origine de cette petite taille de la zone de mélange à Kibuye, pourtant la profondeur maximale du lac est plus large à Kibuye qu'à Bukavu. Ainsi, la plus grande quantité des nutriments reste enfuie en profondeur et ne remonte jamais à la surface où elle pourrait être exploitée par le plancton et influencer la transparence de l'eau. Le zooplancton pélagique du lac Kivu, bassin de Kibuye, connaît des fluctuations mensuelles variables suivant la disponibilité de la nourriture. La plus forte densité a été observée en octobre puisque les vents forts ont été plus intenses en septembre. En 1959, KISS a dit que le zooplancton du lac Kivu subit des fluctuations saisonnières qui atteignent un pic vers août - septembre après le brassage des eaux pendant la grande saison sèche. MAHY (1979) signale que le zooplancton du lac Kivu augmente deux fois par an : après le brassage de la grande et de la petite saison sèche respectivement. En réalité, on peut dire que l'abondance du zooplancton du lac Kivu suit le passage des vents forts qui provoquent le brassage. Nous avons observé la plus grande densité du zooplancton au début d'octobre ce qui suit le pic des vents forts observés en septembre. La plus faible densité observée au mois d'aoüt pourrait être justifiée par la diminution de la biomasse phytoplanctonique (fig. XVII). Concernant la distribution verticale du zooplancton
pélagique au bassin de Kibuye ; c'est la température et la lumière et les paramètres biologiques tel que le phytoplancton. Cette distribution ne dépasse jamais la limite de l'oxygène dissous (60m), puisque le zooplancton est aérobie. FOURNIRET, 1992, ISUMBIHO et al., 2006 et KANINGINI et al. 2003 ont trouvé la même chose dans la partie nord du lac, à Ishungu et à Bukavu respectivement. La couche superficielle remplit toutes les conditions nécessaires à la vie du zooplancton; dans cette couche se trouvent la zone de mélange et la zone euphotique ainsi le phytoplancton trouve les nutriments et réalise la photosynthèse, l'oxygène dissous y est plus concentré que dans d'autres couches, le pH est légèrement basique, la température est favorable etc. Les copépodes dominent d'autres groupes du zooplancton au lac Kivu, puisque l'introduction du L. miodon a entraînée les modifications dans la composition zooplanctonique de ce lac. Il y a eu la disparition du Daphinia curvirostris qui est une espèce de cladocères et la domination numérique du zooplancton par les copépodes. SZLAUER, 1965 et DRENNER et al. 1978 soulignent que les copépodes sont capables de résister à la prédation des poissons planctonophages par leurs mouvements migratoires. STRICKLER, 1997 précise que les copépodes sont favorisés par rapport aux cladocères, puisque le L .miodon est un prédateur visuel. ISUMBISHO et al., (2006) précisent que la domination numérique des copépodes est une caractéristique importante des lacs habités par le L. miodon. Avant l'introduction du L. miodon au lac Kivu, le zooplancton était dominé par les cladocères. Mais après cette introduction, les rotifères ont succédé aux cladocères jusqu'à 1995 oü KANINGINI a trouvé la domination numérique des copépodes. Toute chose étant égale par ailleurs, la maille du filet à plancton utilisé (100um) dans l'échantillonnage pourrait laisser échapper certains rotifères et cladocères de très petite taille. Toutefois, les résultats qu'a trouvé ISUMBISHO et al., 2006 avec un filet à plancton de 55 um confirment la domination numérique des copépodes dans le lac Kivu. Ce qui fait que la faible représentativité des rotifères et des cladocères dans nos échantillons n'a rien à voir avec le vide des mailles du filet à plancton que nous avons utilisé. Néanmoins, nous ne pourrons pas ignorer que la composition spécifique des rotifères pourrait être plus grande si le filet été de plus petite maille. Dans la zone pélagique du bassin de Kibuye, les copépodes sont représentés par 3 espèces qui sont toutes les cyclopoïdes. Les cladocères étaient représentés aussi par 3 espèces. Les rotifères eux étaient représentés par 2 espèces seulement. A Ishungu, ISUMBISHO et al., 2006 ont trouvé aussi 3 espèces de copépodes, 4 espèces de cladocères et 11 espèces de rotifères. Les rotifères pourraient être représentés par beaucoup plus d'espèces si le filet à plancton que nous avons utilisé au bassin de Kibuye n'était pas de 100 um de maille puis que certaines espèces sont de plus petite taille. Au bassin du nord, à l'aide d'un filet de 50 um FOURNIRET, 1992 a trouvé 4 espèces de cladocères, 3 espèces de copépodes et 5 espèces de rotifères. Les crustacés pélagiques du lac Kivu bassin de Kibuye (copépodes et cladocères), migrent vers la surface la nuit et descendent en profondeur le jour. La migration verticale des copépodes adultes n'est pas influencée par le cycle lunaire comme l'a précisé ISUMBISHO et al., 2003. Par contre, les cladocères ne migrent pas de façon significative en période des nuits sans lune. Cette migration verticale des crustacés pélagiques au bassin de Kibuye serait motivée par la lumière solaire, la recherche de la nourriture et l'esquive à la prédation des planctonophages. Les profiles verticales de la conductivité sont les mêmes durant toute la période qu'a couvert notre travail. La structure de la colonne d'eau est influencée par les facteurs météorologiques puisque la profondeur de la zone de mélange (mixolimnion) augmente et atteint 60-70m de profondeur à même temps que le passage des vents forts sur le lac. Lorsque la fréquence des vents forts n'était pas encore plus grande la zone de mélange ne dépassait pas 40m de la profondeur. En réalité, la vitesse des vents forts augmente dans le rift valley pendant la saison sèche. Ainsi, l'augmentation de la vitesse du vent et la diminution de la température de l'eau de surface sont à la base de l'augmentation de la zone de mélange, mais cette dernière ne pourra pas dépasser 70m de profondeur. Au lac Kivu bassin de Kibuye, la chimiocline est stable et sa stabilité résulte d'une importante augmentation de la densité de l'eau à partir de 70m de profondeur comme l'ont signalé [DEGENS et al., 1973 cité par ISUMBISHO et al., 2006]. Au bassin de Kibuye, la stratification verticale de la colonne
d'eau est bien visible sur le profil 52 D'avril à novembre 2005, des échantillons d'eau et de zooplancton ont été prélevés dans la zone pélagique du bassin de Kibuye (partie Est du lac Kivu). Les résultats obtenus indiquent ce qui suit : - Le zooplancton du lac Kivu (Bassin de Kibuye) comprend 3 grands taxa ; les Copépodes, les Cladocères et les Rotifères. - Numériquement, les Copépodes (Thermocyclops consimilis, Mesocyclops aequatorialis, Tropocyclops confinis, copépodites et Nauplii) sont les plus abondants suivis des Cladocères (Diaphanosoma brachyurum, Alona rectangula et Moina micrura) et des Rotifères (Brachionus quadridentatus et Keratella spp.) qui sont les moins abondants. - Le zooplancton est concentré dans les eaux superficielles où les conditions favorables (l'abondance algale, l'oxygène dissous, la température...) à leur survie semblent être réunies. - La plus grande densité du zooplancton pélagique du lac Kivu s'observe jusqu'à une profondeur de 20m et diminue progressivement dans la colonne d'eau jusqu'à 60m de profondeur. - La migration verticale du zooplancton pélagique du lac Kivu bassin de Kibuye n'est pas influencée par la lune, à l'exception des cladocères. L'examen des fluctuations de la densité du zooplancton ont révélé que les périodes de forte abondance sont en relation avec les vents violents qui soufflent sur le lac. Ces périodes de forte abondance se produisent quelques semaines après le passage des vents violents sur le lac dont la périodicité et la fréquence entretiennent les mécanismes de productivité biologique du lac Kivu. Les vents saisonniers paraissent donc être les principaux facteurs qui provoquent une diminution de la transparence des eaux de surface comme c'est le cas dans le lac Tanganyika. A la fin de ce travail nous trouvons que nos hypothèses ont été vérifiées et que nos objectifs ont été atteints. Après cette étude nous recommandons ce qui suit : - Une étude comparative entre le zooplancton de la zone pélagique et celui de la zone littorale permettrait de bien mettre en évidence la distribution horizontale des différentes espèces du zooplancton au lac Kivu, bassin de l'Est. - L'opportunité de poursuivre cette étude sur une longue période en combinaison avec une étude du phytoplancton, de la production primaire, de l'abondance des nutriments et des paramètres physico-chimiques permettrait de bien élucider le fonctionnement biologique du lac Kivu. - Une étude génétique visant l'identification plus précise des espèces du zooplancton du lac Kivu s'avère aussi nécessaire. - Une étude de l'abondance relative et de la migration verticale de chaque espèce s'avère aussi nécessaire. 54
|
|