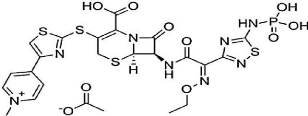
Antibiorésistance des entérobacteries productrices de b-lactamases à spectre étendu isolées de l'hôpital de Sétif (Algérie): détermination des gènes par PCR-multiplexpar Khalil Abdallah REFOUFI Université Ferat Abbas de Setif - Master en Biologie spécialité Microbiologie Appliquée 2016 |

Introductionétroitement associés à des infections nosocomiales. Actuellement, un nouveau type d'enzyme, le CTX-M dissémine au sein des souches communautaires. Les gènes blaTEM, blaSHV, blaCTX-M, blaOXA ont été décrits dans plusieurs études épidémiologiques. La résistance bactérienne par production de BLSE est devenue un problème de santé publique mondiale car compromettant gravement les thérapies antimicrobiennes. De ce fait, le but de ce travail est l'évaluation de la résistance aux f3-lactamines, des souches d'entérobactéries d'isolement courant dans le CHU de Sétif, afin de déterminer la fréquence de résistance aux céphalosporines de 3éme génération et des résistances associées au sein des espèces. Par la suite, une caractérisation par la technique moléculaire (PCR) des f3-lactamases dessouches d'entérobactéries productrices pour mettre en évidence le degré de leur diversité génétique. CHAPITRE 1SYNTHESE BIBLIOGRAPHIQUE 3 Chpaitr 1 : Synthese bibliographique 1- Les entérobactéries 1-1- Définition Les entérobactéries sont des bactéries commensales ou pathologiques du tube digestif de l'homme et des animaux, d'où leur nom (entéro= intestin grêle). Elles peuvent être transitoirement présentes, en dehors du tube digestif, sur différentes parties du revêtement cutanéo-muqueux (Nauciel et Vildé. 2005) et également dans l'environnement. Certaines sont des pathogènes obligatoires provoquent dès leur entrée dans le tube digestif des malades. D'autres, présentent un caractère opportuniste, elles ne sont pathogènes que sur fond d'immunodépression. D'autres groupes prolifèrent dans l'environnement et peuvent être une source de contamination (Grosjean et al. 2011). 1-2- Classification La position phylogénétique de la famille des Enterobacteriaceae se trouve dans l'ordre des Enterobacteriales de la classe des Gammaproteobacteria. Cette famille comprend 176 espèces nommées ou plus, répertoriées en 44 genres (Garrity et al. 2005), dont les plus récents sont Alterococcus, Arsenophorus, Brenneria, Pectobacterium, Raoultella, Samsonia et Sodalis. Les relations phylogénétiques des genres sont fondées sur l'analyse des séquences de l'ARNr 16S des souches types de l'espèce (Garrity et al. 2005).Les genres de cette famille sont regroupés en cinq tribus, d'après leurs propriétés fermentatives : Escherichiae, Klebsielleae, Proteae, Yersiniae et Erwiniae. 1-3- Caractères bactériologiques Les Enterobacteriaceae sont des bacilles à Gram négatif mobiles ou immobiles, avec des extrémités arrondies de 0.5- 1.5 ìm d'épaisseur et 2-4 ìm de longueur, parfois capsulés. Leurs exigences nutritionnelles sont, en général, réduites se développent bien dans un bouillon ou sur gélose ordinaire incubés 18 heures à 37°C. Sur milieu gélosé, ils peuvent donner différentes formes de colonies. Les formes S (smooth) sont l'aspect habituel des colonies lisses, bombés, brillantes et humides. Les formes R (rough) s'observent surtout avec les souches ayant subi plusieurs repiquage où les colonies sont rugueuse, sèches, à contours irréguliers et de teinte mate (Avril et al. 2000). 4 Chpaitr 1 : Synthese bibliographique 1-4- Caractères biochimiques L'identification des Enterobacteriaceae repose sur l'étude des caractères biochimiques. Elles donnent une réaction d'oxydase négative, acidifient le glucose par métabolisme fermentatif, avec ou sans production de gaz et réduisent les nitrates en nitrites. Des galeries biochimiques permettent de déterminer avec précision le genre et l'espèce, se basant sur :
1-5- Caractères antigéniques L'identification des antigènes et sérogroupes a permis de différencier les souches pathogènes des souches commensales. En effet, certains sérotypes ne sont jamais, ou rarement, associés à des maladies tandis que d'autres le sont très fréquemment. L'antigène somatique O, définissant le sérogroupe, est contenu dans les lipopolysaccharides présents sur la paroi bactérienne. L'antigène flagellaire H est de nature protéique entrant dans la structure du flagelle permettant la mobilité de la bactérie. L'antigène de surface K est présent de façon inconstante il bloque l'agglutinabilité de l'antigène O qui peut être restituée après chauffage de la souche à 100°C car il est détruit par ébullition. Les antigènes de surface aussi appelés antigènes de capsule ou d'enveloppe sont appelés « Vi » chez Salmonella (Avril et al. 2000). 1-6- Pouvoir Pathogène Les entérobactéries occupent une place très importante en pathologie humaine infectieuse. Cette importance s'explique aussi bien par la variété des espèces bactériennes qui les composent qu'à leur incidence au niveau de la santé des populations. Les Enterobacteriaceae 5 Chpaitr 1 : Synthese bibliographique forment une vaste famille de bactéries qui sont à l'origine de maladies de gravité très variable, en raison de mécanismes pathogéniques distincts. La fréquence et la gravité des infections, dont elles sont responsables (septicémies, infections nosocomiales, méningites...), traduisent des difficultés de prise en charge liées entre autres à des difficultés d'identification et à leur résistance aux antibiotiques. On les rencontre dans les prélèvements d'origine divers, mais particulièrement dans les urines et les prélèvements sanguins qui constituent une part très importante des activités du laboratoire de bactériologie. Les entérobactéries constituent plus de 80% des germes isolés en laboratoire : Escherichia, Salmonella, Shigella, Klebsiella, Enterobacter, Serratia, Proteus, Morganella et Yersinia sont les bactéries les plus souvent retrouvés. Escherichia coli est un germe très courant. Son habitat est le colon humain où il est le plus abondant. Chez l'homme, la colonisation par E. coli est précoce, et peut être responsable d'un nombre varié de pathologie. Toutefois, trois types de syndromes majeurs résultent de l'infection par de souches E. coli pathogènes : - Infections urinaires (impliqué dans 80 % des infections urinaires), l'incidence de ces infections est plus marquée chez les personnes de sexe féminin en milieu extrahospitalier en raison notamment de la colonisation de la région péri-urétrale. En milieu hospitalier, l'incidence est égale entre les deux sexes en rapport essentiellement avec l'utilisation fréquente des sondes urinaires. - Infections digestives (diarrhées, infections hépatobiliaires et autres), E. coli cause principalement des infections du tractus digestif, en raison de la contamination de l'eau ou des aliments par la flore fécale des malades ou des porteurs. E. coli est subdivisé en 6 pathovars majeurs selon le type de maladie engendrée et les facteurs de virulence associés : les ETEC (E. coli Entérotoxinogènes), les EHEC (E. coli Entérohémorragiques), les EPEC (E. coli Entéropathogènes), les DAEC (E. coli à Adhérence Diffuse), les EAEC (E. coli Entéroagrégatives) et les EIEC (E. coli Entéroinvasives) (Avril et al.2000). - Méningites néonatales et septicémies, E. coli à l'origine de maladies extra- intestinales, a acquis la capacité de déjouer les défenses immunitaires de l'hôte, et à se propager dans l'organisme et il peut coloniser les voies génitales et générer des méningites néonatales. Klebsiella pneumoniae habite le tractus digestif et le système respiratoire supérieur. Ce germe est principalement isolé en milieu hospitalier, le portage étant fortement accru chez les 6 Chpaitr 1 : Synthese bibliographique patients hospitalisés. Toutefois, il est également présent en dehors des hôpitaux, notamment chez des patients diabétiques, fortement débilités, ou souffrant de maladies respiratoires chroniques. Klebsiella pneumoniae peut causer des pneumonies lobaires, des bronchites et broncho-pneumonies, la contamination pulmonaire se faisant surtout par voie aérienne, mais la voie hématogène n'étant pas exclue. Klebsiella pneumoniae est également retrouvé dans des infections urinaires suite au passage de la flore fécale aux voies urinaires. Finalement, des bactériémies compliquent parfois les infections localisées. Klebsiella oxytoca est une bactérie isolée dans la majorité des cas dans les selles, mais peut aussi être isolée dans les urines, le sang et les sécrétions naso-pharyngées et trachéales. A l'instar de Klebsiella pneumoniae, Klebsiella oxytoca peut infecter les voies urinaires et respiratoires des patients hospitalisés. Salmonella constitue un vaste groupe de bactéries, composé de plus de 2000 variétés, et subdivisé en deux sous-groupes. Le premier comprend les Salmonella dites majeures qui causent les fièvres typhoïdes et paratyphoïdes dues à la transmission féco-orale, l'eau ou la nourriture étant contaminées par les selles d'un malade ou d'un porteur sain. Le second est composé des Salmonella dites mineures qui sont à l'origine d'intoxications alimentaires (aliments provenant d'animaux infectés ou contaminés ; viande, produits laitiers...), de gastro-entérites et de bactériémies. Enterobacter cloacae est un germe colonisant souvent les patients hospitalisés et plus particulièrement ceux traités par antibiotiques. Il a été associé à des épidémies nosocomiales et est considéré comme pathogène opportuniste (Hart. 2006). Il peut causer de nombreux types d'infections, y compris abcès cérébraux, pneumonie, méningite, septicémie et infection de plaies, infection des voies urinaires et des infections de la cavité abdominale ou des intestins (Farmer et al. 2007). De plus, il a été observé dans des infections liées à des appareils intra-vasculaires et des infections au point de chirurgie (surtout des infections postopératoires ou liées à des dispositifs comme des prothèses biliaires). Serratia marcescens colonise les systèmes respiratoires, digestifs et urinaires des patients. Bien que les germes isolés en milieu hospitalier soient le plus souvent responsables d'une colonisation asymptomatique, quelques infections nosocomiales sont relatées dans la 7 Chpaitr 1 : Synthese bibliographique littérature: ce sont essentiellement des bactériémies, des infections des voies respiratoires inférieures, des infections urinaires et cutanées. Ces infections sont dues à des germes qui proviennent des patients eux-mêmes ou de leur environnement (Mirabaud. 2003). Citrobacter freundii est un agent pathogène nosocomial opportuniste rare, qui entraîne normalement des infections des voies urinaires, des bactériémies, des sepsis abdominaux et des abcès cérébraux ainsi que des pneumonies et d'autres infections néonatales (Pepperell et al. 2002). 2- Les â-lactamines Les f3-lactamines demeurent les antibiotiques les plus utilisés dans la pratique clinique courante. Cette large utilisation est principalement liée à leur faible toxicité, à leur pouvoir bactéricide et à la diversité des molécules (Robin et al. 2012). L'histoire des B-lactamines débute depuis l'introduction de la pénicilline G en thérapeutique dans les années 1940.Il faudra attendre le début des années 1960 pour voir apparaître les premières synthèses de B-lactamines permettant leur développement à l'échelle industrielle. Un grand nombre de molécules incluant les pénicillines, les céphalosporines, les monobactames et les carbapénémes a été développé. Cependant, la grande utilisation des B-lactamines depuis plus de 60 ans s'est accompagnée d'une augmentation importante de la résistance bactérienne à ces antibiotiques. D'autre part, le développement de nouveaux antibiotiques connaît un franc ralentissement depuis plus de 10 ans (Robin et al. 2012). Cette famille est caractérisée par la présence constante de cycle f3-lactame associé à des cycles et des chaînes latérales variables expliquant les propriétés pharmacocinétiques et le spectre d'activité des différents produits (Cavallo et al. 2004). 2-1- Classification des â-lactamines Quatre groupes de cette classe d'antibiotique peuvent être décrits et ce en fonction de leurs caractéristiques structurales : les pénames (pénicillines), les céphèmes (céphalosporines), les pénèmes et les monobactames (Fig.1).
Chpaitr 1 : Synthese bibliographique
8 Figure 1. Structures chimiques des principales
f3-lactamines et inhibiteurs de f3-lactamases 2-1-1- Les pénicillines (noyau péname) La formule générale des pénicillines associe un noyau B-lactame à un cycle thiazolidine et une chaîne latérale en C-6 (Fig.1). Selon la nature de la chaîne latérale reliée à cette molécule, on distingue plusieurs sous-classes de pénicillines(Tab.1) (Gazengel. 2007). 2-1-2- Les céphalosporines (noyau céphème) Les céphalosporines se distinguent chimiquement des pénicillines par le remplacement du cycle thiazolidine par un cycle dihydrothiazine (noyau « céphème ») avec un atome de soufre 9 Chpaitr 1 : Synthese bibliographique en position 1 (Fig.1). Ces composés sont obtenus par synthèse ou semi-synthèse, à partir de la céphalosporine C, qui est un produit de fermentation d'un champignon (Cephalosporium acremonium), grand producteur de l'acide amino-7-céphalosporanique (noyau de base) (Gazengel. 2007). On distingue cinq générations de céphalosporines (Tab.1). Elles sont classées en fonction de leur date d'apparition, qui correspond à chaque fois à l'acquisition de nouvelles propriétés : ? Les céphalosporines de 1ère génération (C1G) Ont un niveau d'activité assez limité vis-à-vis des bacilles à Gram (-), en raison de leur sensibilité aux f3-lactamases (Nauciel et Vildé. 2005). ? Les céphalosporines de 2éme génération (G) Caractérisées par une meilleure résistance aux B-lactamases et un spectre d'action plus large. Elles possèdent un pouvoir antibactérien un peu plus élevé (CMI plus basses) et un spectre plus large que le 1er groupe vers les entérobactéries (Gazengel. 2007). ? Les céphalosporines de 3éme génération (C3G) L'ensemble de leurs propriétés antibactériennes et pharmacocinétiques autorise des posologies réduites pour une efficacité accrue. Les C3G sont inactives, à des degrés divers par les f3-lactamases à spectre élargi (Nauciel et Vildé. 2005). ? Les céphalosporines de 4éme génération (C4G) Représentées par Céfépime et Céfpirome, elles restent actives chez les entérobactéries ayant acquis une résistance aux C3G par hyperproduction d'une céphalosporinase, et inactives en cas de f3-lactamase à spectre étendu. Elles sont une substitution possible aux céphalosporines de troisième génération pour le traitement de germes résistants (Hardman et Limbird. 1998). ? Les céphalosporines de 5éme génération (C5G) Représentées par la Ceftaroline, Ceftolozane et Ceftobiprole. Ces antibiotiques sont à usage hospitalier. La Ceftaroline a un large spectre d'activité contre de nombreuses bactéries à Gram négatif (Kenneth et al. 2017). 10 Chpaitr 1 : Synthese bibliographique
Figure 2. Structure chimique de la ceftaroline (Jacquline et Tattevin. 2016). 2-1-3- Les pénèmes (Carbapénèmes) Les Carbapénèmes sont des f3-lactamines présentant un très large spectre d'activité et une grande stabilité vis à vis de la plupart des ß-lactamases. L'imipenème et le méropeneme ont été les deux premiers antibiotiques disponibles en clinique. Une troisième molécule s'est ajoutée, l'ertapénème. Parmi les nouvelles Carbapénèmes, le doripénème garde une meilleure activité sur les bacilles à Gram négatif. Les Carbapénèmes sont historiquement considérées comme le traitement de choix des infections sévères à bactéries Gram négatives (Grall et Muller-Serieys. 2013). 2-1-4- Les monobactames L'aztréonam fut le premier monobactame fabriqué synthétiquement, il est réservé au traitement parentéral des infections à bactéries Gram (-). Son spectre d'activité est voisin de celui des C3G (bonne stabilité vis-à-vis des f3-lactamases) (Gazengel. 2007). 2-1-5- Les inhibiteurs des â-lactamases Ces substances n'ont pas une grande activité bactéricide mais ils sont capables de protéger les f3-lactamines en inhibant les pénicillinases. Ce groupe est composé de : ? L'acide clavulanique, utilisé en association fixe avec l'amoxicilline (Augmentin®) ou la ticarcilline (Claventin®). ? Le sulbactam (Bétamaze®), éventuellement en association avec l'ampicilline (sulbactam 500 mg + ampicilline 1 g (Unacim®). ? Le tazobactam associé à la pipéracilline (Tazocilline®) (Gazengel. 2007). ? L'avibactam, utilisé en association avec ceftazidime, ceftaroline et aztreonam. L'Avibactam (NXL104) est un non-f3-lactame inhibiteur de BLSE avec activité contre la plupart des classe A et classe C f3-lactamases ainsi que certaines enzymes de classe D Chpaitr 1 : Synthese bibliographique (Buyck et al. 2016). Actuellement, l'avibactam est en développement dans de nombreux essais cliniques. Tableau 1. Classification des â-lactamines. Noyau Sous-famille Molécules Pénames Pénicillines G et V pénicilline G Pénicillines A ampicilline amoxicilline Pénicillines M oxacilline , cloxacilline Carboxypénicillines Uréidopénicillines ticarcilline, temocilline mezlocilline, Pénémes Carbapénèmes imipéneme méropéneme ertapéneme doripénéme Monobactames Monobactame aztréonam
céfadroxil, céfaclor, céfatrizine, céfradine
11 12 Chpaitr 1 : Synthese bibliographique céfixime, cefpodoxime proxétyl, céfotiam hexétil
2-2- Mode d'action des â-lactamines Les f3-lactamines ciblent les protéines de la membrane cytoplasmique qu'on appelle les PLP (protéines de liaison aux pénicillines) ou PBP (Penicillin Binding Proteins). Ces PLP sont en effet les enzymes impliqués dans la synthèse du peptidoglycane (les transpeptidases, les carboxypeptidases et les transglycosylases). Les f3-lactamines présentent une analogie structurale avec un constituant du peptidoglycane en formation, le dipeptide D-ala-D-ala qui est le substrat naturel de ces enzymes. Elles agissent en « substrat suicide » en bloquant le fonctionnement de ces enzymes inhibant ainsi la formation du peptidoglycane. Par conséquent, cette inhibition entraîne un arrêt de la croissance bactérienne (bactériostase). Le blocage de la multiplication est suivi d'une lyse bactérienne. Le peptidoglycane est dégradé sous l'action d'autolysines, ce qui entraîne finalement la lyse (bactéricide). Le nombre et la nature des PLP varient selon les espèces bactériennes, donc l'affinité des divers PLP n'est pas la même pour toutes les molécules des f3-lactamines. L'effet d'une f3-lactamine est en fonction de son affinité pour les différentes PLP de la bactérie considérée (Gaudy et Buxeraud. 2005). 3- Résistance des entérobactéries aux â-lactamines La résistance bactérienne aux antibiotiques est un véritable problème de santé publique. Depuis l'utilisation des antibiotiques, la résistance n'a jamais cessé d'augmenter et les bactéries ont toujours réussi à contourner les antibiotiques destinés à les tuer. Jusqu'à maintenant, cette évolution était compensée par la mise sur le marché de nouveaux antibiotiques, mais celle-ci s'est considérablement amenuisée laissant le risque de voir apparaître une aire dite « post-antibiotique » (Armand-Lefévre. 2017). 13 Chpaitr 1 : Synthese bibliographique 3-1- Types de résistance La bactérie est considérée résistante lorsqu'elle supporte une concentration d'antibiotique nettement plus élevée que celle qui inhibe la multiplication de la majorité des autres bactéries de la même espèce (Boulahbal. 2010) ou de la tuer (Poole. 2004 ; Schwarz et Chaslus-Dancla. 2001). Les conditions d'activité d'un antibiotique sont de posséder une cible spécifique, de demeurer sous forme active, d'accéder à la cible et d'interagir efficacement avec elle en la désactivant. Les entérobactéries sont soit sensibles aux â-lactamines, soit naturellement résistantes ou soit ont une résistance acquise (Vora et Auckenthaler. 2009). 3-1-1- La résistance naturelle La résistance naturelle est programmée sur le génome et constante à l'intérieur du taxon ; elle constitue un critère d'identification stable d'une espèce. La résistance naturelle ou intrinsèque à un antibiotique est essentiellement due à la présence de gènes chromosomiques; elle est donc commune à toutes les bactéries d'une même espèce et transmise à la descendance. Dans tous les cas, la résistance naturelle fait partie des caractères normaux de l'espèce ; elle détermine le niveau de sensibilité « basal » des bactéries et définit le phénotype sauvage d'une espèce. Elle est constitutive et touche toute une famille d'antibiotiques (Mayer et al. 2000). Ainsi, les entérobactéries ont une résistance naturelle à la pénicilline G, V et les pénicillines du groupe M (Philippon et Arlet. 2012). 3-1-2- La résistance acquise Ce terme est utilisé pour désigner le résultat d'un processus permettant à des bactéries d'une espèce originellement sensible de devenir résistante à un ou plusieurs antibiotiques. L'acquisition de ces résistances est déterminée par des modifications génétiques consécutives à des mutations ponctuelles (plutôt rare) ou à l'acquisition de nouveaux gènes de résistance exogènes (plutôt fréquent) par le biais de plasmides, de bactériophages, de transposons ou d'intégrons (Davies. 1997 ; Chopra et al. 2003). La capacité de multiplication très rapide des bactéries favorise la sélection d'évènements génétiques favorables et la possibilité d'échange d'information même entre espèces lointaines leur conférant un très grand pouvoir d'adaptation aux contraintes du milieu. L'évolution des 14 Chpaitr 1 : Synthese bibliographique mécanismes de résistance aux antibiotiques, et notamment aux â-lactamines illustre parfaitement ce phénomène. 3-2- Supports génétiques de la résistance Parmi les modalités d'acquisition de la résistance aux antibiotiques, le transfert horizontal de gènes est un élément-clé. Ce type de transfert a probablement lieu dans tous les écosystèmes terrestres colonisés par les bactéries. Ainsi ces transferts ont été mis en évidence dans de nombreux écosystèmes, tels les sols, les rivières, les environnements marins, mais aussi les tubes digestifs d'insectes ou de mammifères. Différents éléments génétiques sont impliqués dans ce transfert de gènes. 3-2-1- Le chromosome La résistance chromosomique acquise résulte d'une mutation. La fréquence de ces mutations est faible et variable (10-6 à 10-9) mais la mutation est stable et transmissible à la descendance. La résistance apparaît dans ce cas au hasard et n'est donc pas influencée par l'antibiotique quine fait que la révéler. 3-2-2- Les éléments génétiques mobiles La résistance aux antibiotiques acquise par les bactéries est principalement due à la présence de trois types d'éléments extra-chromosomiques portant des gènes de résistance : les plasmides, les transposons et des cassettes de résistances insérées sur un intégrons (Bennett. 2008 ; Martinez. 2010 ; Walsh. 2006). Ce type de résistance est transmissible d'une souche ou d'une espèce à une autre selon trois modes de transmission, à savoir la conjugaison (transfert direct entre deux bactéries ayant établi temporairement un contact physique), la transformation (transfert d'ADN nu) et la transduction (transport d'ADN bactérien par des bactériophages). 15 Chpaitr 1 : Synthese bibliographique
Figure 3. Modes de transmission du matériel génétique : la conjugaison, la transformation et la transduction chez les bactéries (Andersson et Hughes. 2010). a) Plasmides Les plasmides bactériens revêtent une importance toute particulière dans l'étude des phénomènes de résistance car ils constituent à la fois un vecteur de premier plan pour la dissémination des résistances et un immense réservoir génétique. Comme les chromosomes, les plasmides codent pour des protéines et des molécules d'ARN et se répliquent durant la croissance bactérienne. Les plasmides partagent certaines fonctions de partition et quelques recombinasses spécifiques de site avec le chromosome de l'hôte (Snyder et Champness. 2007). Contrairement aux chromosomes, les plasmides ne portent toutefois pas de gènes essentiels à la croissance bactérienne. Ils codent plutôt pour des protéines dont la fonction est de conférer un avantage sélectif comme la résistance aux antibiotiques ou encore des facteurs de virulence. Les plasmides peuvent conférer une résistance aux principales classes d'antibiotiques (Carattoli. 2009). Une même bactérie peut porter un ou plusieurs plasmides selon leur compatibilité mutuelle et chacun d'eux peut contenir un ou plusieurs gènes de résistance (Harbottle et al. 2006).Les plasmides peuvent également se transmettre à une souche sensible principalement par conjugaison mais aussi par transformation ou transduction à l'aide de bactériophages. Les plasmides, en fonction de leur efficacité de conjugaison, vont permettre le 16 Chpaitr 1 : Synthese bibliographique transfert horizontal de ces composantes à d'autres bactéries qui peuvent être de différentes espèces, genres et familles (Thomas et Nielsen. 2005).Ils peuvent acquérir d'autres gènes de résistance par l'intermédiaire d'éléments transposables et d'intégrons par exemple. La survenue épidémique de bactéries à Gram négatif multirésistantes est largement attribuée à ce type de transmission.
Les intégrons, plus récemment décrits vers la fin des années 80, jouent un rôle majeur dans l'acquisition et l'expression de gènes de résistance aux antibiotiques, notamment chez les bactéries à Gram négatif. Le rôle d'une partie de ces intégrons, parfois nommés intégrons de résistance (IR), est majeur dans la dissémination de la résistance aux antibiotiques et notamment chez des souches d'intérêt clinique. L'ampleur de la diffusion de la résistance est attribuable en majeure partie au déplacement des gènes de résistance par le biais d'intégrons (Domingues et al. 2012).Ceux-ci sont des éléments génétiques mobiles avec une structure spécifique composée de deux segments conservés encadrant une région centrale appelée «cassette » (Domingues et al. 2012 ; Recchla et Hall. 1997). La transposition des intégrons n'est pas anarchique et a lieu au niveau de sites spécifiques, celle-ci peut recevoir, a différents moment des gènes variés dont ceux de la résistance aux antibiotiques qui vont s'insérer toujours au même endroit « site d'insertion » ce qui provoque l'allongement de l'intégrons 17 Chpaitr 1 : Synthese bibliographique dans un sens. Les intégrons peuvent être associés à des (SI), des transposons ou des plasmides pouvant leur conférer une mobilité (Domingues et al. 2012). 3-3- Mécanismes de résistance aux â-lactamines Les différents mécanismes de résistance aux f3-lactamines peuvent s'exprimer seuls ou de façon concomitante agissant alors de façon synergique. Il peut s'agir d'une diminution d'affinité de la cible, d'une diminution de la perméabilité de la membrane, de l'expression d'une pompe d'efflux actif ou de la production d'enzyme de type f3-lactamase (Vora et Auckenthaler. 2009). 3-3-1- Diminution de la perméabilité La pénétration des f3-lactamines, à travers la membrane externe s'effectue à travers les porines qui sont des canaux protéiques remplis d'eau. Ainsi, la sensibilité aux f3-lactamines dépend du nombre de porines fonctionnelles. L'altération des porines par mutation est à l'origine de résistances acquises aux f3-lactamines, soit par une modification structurale d'une porine essentielle, ce qui a été décrit chez E. coli, soit par une diminution quantitative des porines, qui est la situation la plus fréquente (Kumar et Schweizer. 2005). 3-3-2- Modification de la cible La résistance aux f3-lactamines, conférée par les PLPs, chez les bactéries à Gram négatif joue un rôle mineur dans la résistance (Spratt et al. 1989). Cette résistance peut avoir lieu par des mutations dans les gènes chromosomiques codant pour les PLPs ou par l'acquisition de gènes étrangers codant pour de nouveaux PLPs ayant une affinité différente aux f3-lactamines (Georgopapadakou. 1993). Une diminution de la production de la PLP1A a été associée à la résistance pour l'imipénème et au mécillinam chez P. mirabilis suite à une perte d'affinité de la PLP2 et à une diminution de la quantité de PLP1 (Neuwirth et al. 1995).Ces mutations chez les entérobactéries restent rares (Robin et al. 2012). 3-3-3- Hyperproduction du système d'efflux Les systèmes d'efflux éliminent activement les f3-lactamines dans le milieu extérieur en utilisant la force proton-motrice. Ainsi, ils diminuent la concentration d'antibiotique au contact de sa cible et donc son efficacité. Le système d'efflux actif est efficace grâce aux Chpaitr 1 : Synthese bibliographique protéines transmembranaires ancrées dans la membrane cytoplasmique mais également dans la membrane externe des bactéries à Gram négatif (Walsh. 2003). L'implication des systèmes d'efflux dans la résistance aux f3-lactamines a été clairement identifiée dans plusieurs études en particulier chez K. pneumoniae. Cependant ce type de mécanisme touchant préférentiellement la céfoxitine et les G semble difficile à distinguer du point de vue phénotypique des résistances par modification des porines (Bialek-Davenet et al. 2011 ; Robin et al. 2012). 3-3-4- Production d'enzymes La production des f3-lactamases est le principal mécanisme de résistance des entérobactéries aux f3-lactamines (Abbas et al. 2012).Cependant, toutes les entérobactéries, quel que soit leur groupe, sont capables d'intégrer des gènes de résistance codant pour une f3-lactamase (Zoghleb et Dupont. 2005). 4- Les â-lactamases C'est le mécanisme qui nous intéresse le plus dans notre étude. Les f3-lactamases sont des enzymes bactériennes hydrolysant la fonction amine du cycle f3-lactame et rendant l'antibiotique inactif avant qu'il n'atteigne sa cible : les PLP. Les f3-lactamases sont des enzymes constitutionnelles ou acquises, produites par les bactéries. Leur activité enzymatique provoque l'ouverture du cycle f3-lactame et crée un intermédiaire acyl-enzyme instable qui est ensuite dégradé en acide inactif (Vodovara et al. 2013).Les gènes codant pour ces enzymes sont principalement situés sur des éléments génétiques mobiles (plasmides, transposons, intégrons) (Boyer et al. 2011).
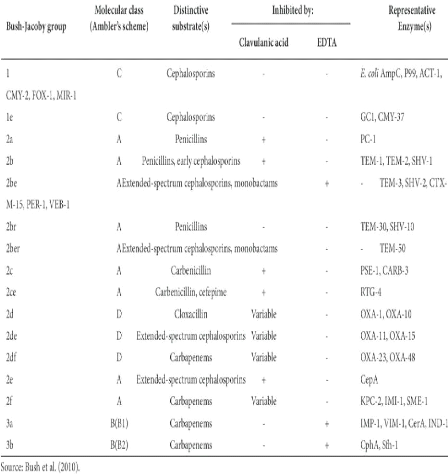
18 Figure 4. Mode d'action des f3-lactamases (Barrial et Scotet. 2006). 19 Chpaitr 1 : Synthese bibliographique 4-1- Classification des â-lactamases Les deux classifications couramment utilisées sont celle d'Ambler et de Bush, Jacoby et Medeiros. La classification structurale d'Ambler est basée sur la séquence peptidique du site enzymatique et distingue quatre classes, alors que la classification fonctionnelle de Bush et ses collaborateurs repose sur l'activité hydrolytique et la sensibilité des f3-lactamases aux inhibiteurs. Elle rend compte de leur diversité fonctionnelle au sein des quatre classes structurales d'Ambler, notamment dans sa classe A (Vodovara et al. 2013).Aujourd'hui, les classifications d'Ambler et de Bush-Jacoby-Medeiros sont considérées comme étant les plus pertinentes (Bush et al. 1995). 4-1-1- Classification d'Ambler ? Les â-lactamases de classe A Les f3-lactamases de classe A, d'origine chromosomique ou plasmidique, se caractérisent par leur capacité à hydrolyser l'amide cyclique lié à la molécule de f3-lactame. Cette activité hydrolytique, assurée par une sérine conservée (Ser-70)- induit la formation d'acide pénicilloïque pour la pénicilline et de céphalosporoate, analogue aux pénicillinoates, pour les céphalosporines. Cette sérine est retrouvée dans le site actif de la f3-lactamase et assure un fort taux de résistance aux pénicillines, céphalosporines et carbénicillines. Cependant, les f3-lactamases de classe A sont sensibles aux inhibiteurs de f3-lactamases (Livermore. 1995). ? Les â-lactamases de la classe B Les f3-lactamases de classe B sont des métallo-f3-lactamases et utilisent un ion de zinc (Zn2+) comme cofacteur permettant ainsi la décomposition de l'anneau f3-lactame (Ambler. 1980 ; Bandoh et al. 1991). Ces enzymes ont émergés depuis une dizaine d'années, d'abord chez Pseudomonas aeruginosa et Acinetobacter baumannii, puis chez les entérobactéries et d'autres bacilles à Gram négatif. L'importance clinique des métallo-bêta-lactamases est liée au fait qu'elles hydrolysent les carbapenèmes, composés qui échappent à l'activité des f3-lactamases à sérine active. La plupart des métallo-f3-lactamases hydrolysent une variété de pénicillines et de céphalosporines, et sont insensibles aux inhibiteurs suicides classiques (Bebrone. 2007). 20 Chpaitr 1 : Synthese bibliographique ? Les â-lactamases de la classe C Les f3-lactamases de classe C, décrites en 1981 par Jaurin et Grundstrom, ont longtemps été ubiquitaires aux chromosomes de différentes espèces bactériennes entériques (Jaurin et Grundstrom. 1981).Dans la classe C, on retrouve les céphalosporinases AmpC qui sont codées par des gènes qui étaient primitivement situés sur le chromosome de nombreuses bactéries à Gram négatif telles que Citrobacter freundii, Serratia marcescens et Enterobacter spp. Chez ces bactéries, l'expression d'AmpC est inductible, les gènes codant pour ces enzymes sont aussi présents chez E. coli, où ils ne sont pas inductibles. Ces enzymes sont résistantes à l'acide clavulanique (Philippon et al. 2002). ? Les â-lactamases de la classe D Les f3-lactamases de classe D se distinguent par leur capacité à hydrolyser les pénicillines isoxazolyl (oxacilline) et la méthicilline, et sont faiblement inhibées par l'acide clavulanique. Elles sont retrouvées fréquemment chez Pseudomonas aeruginosa et Acinetobacter spp. (OXA-11, 14, 16, 17 et 19) et peu chez les enterobactéries (Paterson et Bonomo. 2005).De plus, certains membres de cette famille, dont OXA-10 (PSE-2) et OXA-11 (Hall et al. 1993), possèdent un large spectre d'activité assurant une forte résistance aux céphalosporines de troisième génération dont la céfotaxime, le céfoperazone et la ceftazidime (Hall et al. 1993 ; Livermore. 1995). 4-1-2- Classification de Bush-Jacoby-Medeiros Elle est établie selon les propriétés fonctionnelles de l'enzyme définies par son substrat préférentiel et son profil d'hydrolyse (Bush et al. 1995). Ces auteurs divisent ces enzymes en quatre groupes (1 à 4) avec plusieurs sous-groupes. Les bêtalactamases de la classe A sont ainsi subdivisées en 8 groupes fonctionnels 2a, 2b, 2be, 2br,2ber, 2c, 2e et 2f (Bush et Jacoby. 2010). Cette classification phénotypique de f3-lactamases pose un certain problème puisque différentes mutations ponctuelles peuvent influencer leur susceptibilité à ces substrats et inhibiteurs (Jacoby et Medeiros. 1991). Un autre aspect restrictif de cette classification relève du fait que l'attribution d'un acronyme à un gène de résistance n'a plus la même signification. Par exemple, les f3- lactamases PSE, ou "Pseudomonas Specific Enzymes", ont aussi été identifiées chez les Enterobacteriaceae, ce qui remet en question la pertinence de ce préfixe (Matthew et al. 1979). 21 Chpaitr 1 : Synthese bibliographique Tableau 2. Classification global des BLSE (Bush et al. 2010).
22 Chpaitr 1 : Synthese bibliographique 5- Les â-lactamases à spectre élargi BLSE Les BLSE ont été décrites pour la première fois en 1983, en Allemagne (Knothe et al. 1983), il n'y a pas de consensus concernant la définition de f3-lactamases à spectre étendu (BLSE). Classiquement, les BLSE sont définies comme des enzymes appartenant à la classe A (à l'exception des BLSE de type OXA classe D de la classification d'Ambler), capables d'hydrolyser les pénicillines, céphalosporines de première, deuxième, troisième et quatrième génération (céfépime ou cefpirome) et l'aztréonam. Elles sont inhibées in vitro par les inhibiteurs des f3-lactamases (acide clavulanique, tazobactam et sulbactam) (Livermore. 1995). Par contre, les BLSE sont sensibles aux céphamycines (céfotétan et cefoxitine) ainsi qu'aux carbapénèmes. Une co-résistance avec les aminosides, les tétracyclines et les fluoroquinolones est fréquente. Les bactéries possédant des BLSE sont dites multirésistantes (Paterson et Bonomo. 2005). Il s'agit d'un mécanisme de résistance de type plasmidique, et donc transmissible à d'autres bactéries. La présence de ce type de mécanisme de résistance au sein de souches pathogènes fait peser un risque majeur d'inadéquation thérapeutique et donc d'échec thérapeutique (Schwaber et Carmeli. 2007), et est également un facteur de diffusion. Au sein des entérobactéries, Klebsiella pneumoniae et Escherichia coli sont les deux espèces les plus fréquemment porteuses de ces mécanismes de résistance. Toutefois, ces enzymes ont été retrouvées au sein de nombreuses autres espèces bactériennes, entérobactéries et bacilles non fermentant (tels que Pseudomonas aeruginosa et Acinetobacter baumannii) (Jacoby et Munoz-Price. 2005). 5-1- Différents types de BLSE Elles sont classées selon leurs types moléculaires, les plus fréquents étant les types TEM, SHV, CTX-M (Jacoby et Munoz-Price. 2005) et les nouvelles étant les carbapénèmases. 5-1-1- BLSE de type TEM (Temoneira - nom du patient) La première f3-lactamase plasmidique de type TEM (TEM-1) a été isolée en 1965, en Grèce, à partir d'une souche d'E. coli isolée chez une patiente nommée Temoneira, d'où la nomination (Datta et Kontomichalou. 1965). La majorité des BLSE de ce type dérivent par quatre à sept mutations ponctuelles de l'enzyme originale (TEM-1 ou TEM-2). Les substitutions les plus courantes sont le glutamate en lysine en position 104, l'arginine en Chpaitr 1 : Synthese bibliographique sérine en position 164, la glycine en sérine en position 238 et le glutamate en lysine en position 240 (Bradford. 2001). Une seule de ces substitutions peut entraîner une importante modification de l'affinité de l'enzyme. Par exemple, lorsque l'acide aminé devient une sérine, l'interaction entre le -NH3 de la sérine et le groupement oxy-amino des C3G se traduit par une meilleure stabilité de la â-lactamine dans le site actif de l'enzyme. Ces mutations rendent l'enzyme capable d'hydrolyser les C3G, mais aussi plus vulnérable à l'action des inhibiteurs (acide clavulanique). Cependant, d'autres mutations peuvent conférer la résistance aux inhibiteurs. Ces variantes sont appelées TRI (TEM résistantes aux inhibiteurs). Les enzymes dérivées par mutations permettant d'hydrolyser à la fois les C3G et les inhibiteurs sont de plus en plus fréquentes (Rodriguez-Villalobos et Struelens. 2006).
Figure 5.Les BLSE dérivées de TEM (Ambler et al. 1991). 23 24 Chpaitr 1 : Synthese bibliographique 5-1-2- BLSE de type SHV : (Sulfhydryl variable) Les enzymes BLSE de type SHV dérivent par mutations ponctuelles de l'enzyme originale SHV-1 qui correspond a un gène blaSHV de pénicillinase chromosomique de K. pneumoniae (Brisse et Verhoef. 2001 ; Haeggman et al. 2004), et cette désignation SHV est liée au sulfhydryl variable. La majorité des BLSE de type SHV est caractérisée par la substitution d'acides aminés Gly238Ser ou Gly238Ser et Glu240Lys. Le résidu sérine à la position 238 est indispensable pour l'hydrolyse efficace du céfotaxime et le résidu lysine est crucial pour l'hydrolyse efficace de la ceftazidime (Elhani. 2012). La majorité des BLSE de type SHV ont été décrite chez les souches de K.pneumoniae, toutefois ces enzymes ont été trouvées chez C. freundii, C. diversus, E. coli, E. cloacae (Bradford. 2001).La présence de la séquence d'insertion IS26 sur le gène SHV faciliterait l'acquisition du phénotype BLSE (Hammond et al. 2005).
Figure 6.Les BLSE dérivées de SHV (Bradford. 1999). 25 Chpaitr 1 : Synthese bibliographique 5-1-3- BLSE de type CTX-M (Cefotaximase-Munich) Les BLSE de type CTX-M ont été décrites initialement en 1986 (FEC-1) au Japon, en Allemagne et en France en 1989 (CTXM-1) et ont depuis lors disséminé largement dans le monde (Thomson et Moland. 2000). Ces enzymes représentent à l'heure actuelle les BLSE les plus fréquentes au niveau mondial après leur diffusion rapide depuis les années 90 (Bonnet. 2004 ; Livermore et al. 2007). Le groupe CTX-M (pour céfotaximase) conférait à l'origine, chez les entérobactéries, un plus haut niveau de résistance au céfotaxime (ou ceftriaxone), céfépime et aztréonam qu'à la ceftazidime (Arlet et Philippon. 2003 ; Bonnet. 2004). Certaines d'entre elles ont évolué plus récemment par mutation (ponctuelle ou non) générant un haut niveau de résistance à la ceftazidime telles les enzymes CTX-M-15, CTX-M-16, CTX-M-19, CTX-M-23 et CTX-M-32 (Bonnet. 2004). Les CTX-M sont plus fortement inhibées par le tazobactam que par l'acide clavulanique. Ces nouvelles BLSE ne sont pas étroitement liées aux â-lactamases de type TEM ou SHV puisqu'elles ne présentent que 40 % d'homologie avec ces BLSE classiques (Elhani. 2012). Les analyses génétiques ont montré que les gènes progéniteurs appartiennent au genre Kluyvera, entérobactéries d'isolement très rare en bactériologie médicale (Bonnet. 2004 ; Humeniuk et al. 2002). La â-lactamase naturelle de Kluyvera cryocrescens (KLUC-1) présente 95 à 100 % d'identité avec les enzymes plasmidique du phylum CTX-M-1 (Decousser et al. 2001). 5-1-4- Autres types de BLSE D'autres BLSE ont une distribution moins large, caractérisées par un haut niveau de résistance à la ceftazidime et parfois à l'aztréonam plutôt qu'au céfotaxime (Arlet et Philippon. 2003 ; Bradford. 2001), qui sont individualisées en BES-1 (brazilian extended spectrum), GES-1 (Guyana extended spectrum), PER-1 (Pseudomonas extended resistance) (Weldhagen et al. 2003), SFO-1 (Serratia fonticola), TLA-1 (Tlahuicas, tribu mexicaine), et VEB-1 (Vietnam extended spectrum). Des enzymes proches de GES-1 ont été découvertes en Grèce, malheureusement dénommées à tort IBC (integron borne cephalosporinase) (IBC-1, IBC-2) (Philippon et Arlet. 2006). Enfin, l'OXA-1 qui a une grande activité catalytique pour la cloxacilline, l'oxacilline et la méticilline. 26 Chpaitr 1 : Synthese bibliographique 5-1-5- Les Carbapénèmases Les Carbapénèmases décrites chez les entérobactéries appartiennent aux quatre classes connues de f3-lactamases (classe A, B, C, D de la classification d'Ambler) (Queenan et Bush, 2007 ; Poirel et al. 2007). Les plus importantes actuellement en clinique sont les KPC, MBL et OXA-48. L'activité des Carbapénèmases de classe A est inhibé in vitro totalement ou partiellement par les inhibiteurs des f3-lactamases (acide clavulanique et tazobactam), alors que les autres carbapénèmases sont insensibles à ces inhibiteurs classiques de f3-lactamases (Nordmann et Carrer. 2010). a) Classification des carbapénèmases ? Les Carbapénèmases de classe A Les carbapénèmases de classe A ont été tout d'abord rapportées dans plusieurs souches d'entérobactéries isolées de l'environnement (Serratia, Enterobacter), produisant des bêta-lactamases dont l'activité était inhibée par l'acide clavulanique. Elles hydrolysaient, à divers degrés, toutes les bêtalactamines. Leurs gènes sont chromosomiques ou plasmidique. Il s'agissait des bêtalactamases NmcA, Sme-1, Sme-2/Sme-3, IMI-1/IMI-2, ou SFC-1. D'autres carbapénémases, de type GES, ont été identifiées chez K. pneumoniae et E. coli. Ces enzymes hydrolysent les carbapénèmes faiblement (Queenan et Bush. 2007 ; Poirel et al. 2007). Les carbapénémases de classe A, les plus fréquentes et les plus menaçantes, sont les carbapénémases de type KPC (Nordmann et al. 2009). La première souche exprimant KPC fut identifiée chez une souche de K. pneumoniae en 1996, en Caroline du Nord aux États-Unis (Yigit et al. 2001). Elle hydrolyse toutes les bêtalactamines. Son activité est partiellement inhibée par l'acide clavulanique ou le tazobactam. Le plus souvent les souches qui produisent KPC expriment également d'autres bêta-lactamases dont de nombreux types de BLSE (TEM, SHV, CTX-M) et possèdent un certain degré de résistance par imperméabilité. Les souches KPC apparaissent donc le plus souvent multirésistantes aux bêta-lactamines, l'ertapénème étant la carbapénème dont le niveau de résistance est le plus élevé. Les études génétiques montrent que ces gènes KPC sont localisés sur une variété importante de plasmides mais qu'ils sont associés à des transposons de même nature de type Tn3. La mobilité de ces plasmides et transposons contribuerait fortement à la diffusion interespèce de ces gènes KPC (Nordmann et al. 2009). L'association des gènes KPC à d'autres gènes de résistance aux 27 Chpaitr 1 : Synthese bibliographique antibiotiques sur de mêmes structures génétiques explique en grande partie la multirésistance de ces souches. Tableau 3.Principales carbapénémases des entérobactéries (Nordmann et al.2012).
28 Chpaitr 1 : Synthese bibliographique ? Les Carbapénèmases de classe B Les premières carbapénémases de classe B (IMP) (ou métallo bêtalactamases, MBL) avaient été identifiées chez des espèces d'entérobactéries typiquement hospitalières (Serratia, Citrobacter et Enterobacter) au Japon (Queenan et Bush. 2007 ; Walsh. 2008). Puis d'autres MBL ont été isolées dans le monde entier ; il s'agit de nombreuses variétés de bêtalactamases de type IMP et VIM, GIM-1, KHM-1 et NDM-1 (Walsh. 2008 ; Yong et al. 2009). Ce sont des métallo-enzymes qui contiennent des ions zinc dans leur site actif. Ces enzymes hydrolysent fortement toutes les bêtalactamines à l'exception de l'aztreonam. Leur activité n'est inhibée ni par l'acide clavulanique ni par le tazobactam. Les niveaux de résistance aux carbapénèmes sont assez variables (Queenan et Bush. 2007 ; Walsh. 2008). Dans de nombreux cas, les souches productrices de MBL produisent aussi des BLSE (Walsh. 2008). Les gènes de ces MBL sont, le plus souvent, plasmidique et associés au sein d'intégrons et de transposons, structures assurant la mobilité de ces gènes de résistance et la multirésistance aux antibiotiques des souches (Queenan et Bush. 2007 ; Walsh. 2008). ? Les Carbapénèmases de classe C et D Les carbapénèmases de classe D, OXA-48, décrites tout d'abord chez K. pneumoniae (Poirel et al. 2004) hydrolysent fortement les carbapénèmes et n'hydrolysent pas les céphalosporines de 3e génération. Son activité n'est pas inhibée par l'acide clavulanique (Poirel et al. 2004) OXA-48 est souvent associée à d'autres bêtalactamases, en particulier des BLSE, ce qui contribue à la multirésistance des souches (Carrer et al. 2010 ; Gülmez et al. 2008). En l'absence d'autres bêtalactamases, les souches qui ne produisent que l'OXA-48 peuvent ne présenter qu'une légère diminution de la sensibilité aux carbapénèmes (Aktas et al. 2008). Le réservoir naturel de ce gène OXA-48 a été identifié ; il s'agit de Shewanella sp., ce qui suggère le transfert de ce gène de résistance en milieu aqueux (Poirel et al. 2004) Le gène d'OXA-48 est localisé au sein d'un transposon comportant deux séquences d'insertion identiques assurant mobilité et expression (Poirel et al. 2004). 29 Chpaitr 1 : Synthese bibliographique 6- Epidémiologie mondiale des entérobactéries BLSE 6-1- Les BLSE classiques Depuis plus de 30 ans, la résistance des entérobactéries aux C3G ne cesse de se renforcer notamment par l'acquisition de BLSE. De nombreuses études relatent la progression continue à l'échelle mondiale de ce type de résistance (Belmonte et al. 2010). L'avènement des BLSE a commencé en Europe de l'Ouest mais a gagné rapidement les Etats-Unis, l'Asie et même l'Afrique. Aujourd'hui leur répartition est mondiale et leur prévalence très variable selon la localisation, l'espèce bactérienne et l'origine des isolats (Bradfor. 2001a). L'épidémiologie des BLSE a été étudiée par de grands programmes de surveillance antimicrobienne notamment le programme SENTRY dans la région Asie-pacifique entre 1998 et 2002 et le programme MYSTIC (Meropenem Yearly Susceptibility Test Information Collection) en Europe et aux Etats-Unis entre 1997 et 2004. 6-1-1- En Europe La première bêta-lactamase a été isolée en Allemagne. Il s'agissait du SHV-2 décrit chez K. pneumoniae en 1983 (Knothe et al. 1983) puis en 1985 chez K. ozaenae (Kliebe et al. 1985). En France, en 1984, une variante de TEM-2, nommée TEM-3 est détectée également chez K. pneumoniae dans différents hôpitaux. En 1989, presque simultanément en Allemagne en France et en Italie, une nouvelle famille de BLSE a été reconnue et nommée CTX-M. En 1991, à Ankara (Turquie), et plus tard en France, les enzymes de type OXA, ont été découvertes (Canton et al. 2008). Les données du programme de surveillance MYSTIC en Europe entre 1997 et 2004 ont montré une augmentation de la prévalence et la dispersion des souches productrices de BLSE. Chez les souches de E. coli productrices de BLSE, la prévalence va de 2,1 % en 1997 à 10,8 % en 2004; chez K. pneumoniae elle va de 9,0 % à 13,6 % (Goossens et Grabein. 2005). 6-1-2- En Afrique Deux études ont permis de faire le point sur la production des BLSE en Afrique. La première relève la diversité de la prévalence des souches productrices de BLSE sur le continent et conclut que la proportion de ces souches est inférieure à 15 % (Tansarli et al. 2014). Dans la partie nord de l'Afrique, on retrouve les gènes CTX-M-3, CTX-M-15 et TEM-1 chez K. 30 Chpaitr 1 : Synthese bibliographique pneumoniae et TEM, SHV et CTX-M-1 chez E. cloacae avec une prévalence de 32,1% parmi les entérobactéries en Algérie (Messai et al. 2008; Ahmed et al. 2012; Nedjai et al. 2013).Au Maroc, la prévalence est de 1.3 % avec les types TEM-1, SHV-5 et CTX-M-15 chez E. coli (Bourjilat et al. 2011). En Ethiopie, la prévalence de Klebsiella spp productrice de BLSE est de 33,3 % (Seid et Asrat. 2005) En Afrique du sud, une étude identifie TEM-1,-26, SHV-2,-5 chez Proteus mirabilis et E. coli (Pitout et al. 1998). En Afrique centrale, il a été rapporté au Cameroun les BLSE de type SHV-12, CTX-M-15, OXA-1, OXA-30, TEM-1 avec un portage fécale de 54,06 % (Lonchel et al. 2013a; Lonchel et al. 2013b).En Centrafrique, la prévalence est de 4 % chez les entérobactéries avec les types SHV-2A, SHV-12, CTX-M-3, CTX-M-15 et TEM-1 (Frank et al. 2006). En Afrique de l'Ouest, E. coli productrice de CTX-M-15 est retrouvé chez les enfants au Sénégal (Ruppé et al. 2009) et où la prévalence est estimée à 3,8 % (Sire et al. 2007). Au Nigeria, CTX-M-15, SHV, TEM ont été signalés chez K. pneumoniae (Soge et al. 2006) et OXA-10, VEB-1, CMY chez Providencia spp (Aibinu et al. 2011). La deuxième étude conclut que les entérobactéries hospitalières et communautaires productrices de BLSE varient entre les pays et selon les échantillons considérés mais reste communes en Afrique. Ce sont les enzymes de classe A et D (Storberg. 2014). La vraie prédominance des BLSE n'est pas connue, probablement en raison des difficultés de leur détection. Mais il est clair que leur prévalence et leur distribution mondiale augmente de jours en jours. 6-1-3- En Asie En Asie, des études sur la résistance aux antibiotiques ont montré des niveaux élevés des phénotypes BLSE en Chine, en Corée, au Japon et en Inde. Des travaux à la fin des années 1990 suggèrent que SHV-5 et SHV-12 étaient les plus communs. En Chine, et d'autres pays d'Asie, les types dominants sont CTX-M-14 et CTX-M-3 et CTX-M-2 au Japon. En Inde la prévalence des phénotypes BLSE chez E. coli et K. pneumoniae est de68 % (Hawkey. 2008). Dans le cadre du programme SENTRY, l'incidence de la production de BLSE chez E. coli varie de 13 % à 35 % et un taux > 20 % chez K. pneumoniae, en Chine, au Japon, en Taïwan, au Singapore (Hirakata et al. 2005). 31 Chpaitr 1 : Synthese bibliographique 6-1-4- En Amérique Aux Etats-Unis, les grandes études de surveillance ont été menées par le CDC et par le programme MYSTIC. L'occurrence de la production de BLSE chez les entérobactéries s'étend de 0 à 25%, selon les établissements, avec une moyenne autour de 3% (CDC National Nosocomial Infections Surveillance http://www.cdc.gov/ncidod/hip/SURVEILL/NNIS.HTM et Bradford, 2001a). Une analyse des données du programme MYSTIC entre 1999-2004 aux Etats-Unis a révélé 5,1 % et 7,2 % de phénotype BLSE chez les souches de E. coli et Klebsiella spp, respectivement en 1999 et une baisse en 2004, notamment 1,4 % pour E. coli et 4,4 % pour Klebsiella spp (Goossens et Grabein. 2005). Peu de CTX-M ont été signalés aux Etats-Unis, contrairement au reste du monde (Bush. 2008). Au Canada, une grande variété de BLSE a été identifiée, avec des membres de la classe TEM, SHV et CTX-M-14. Au Mexique l'enzyme TLA-1 hébergé par une souche d'E. coli a été rapportée en 1993 (Naas et al. 2008). En Amérique du Sud le taux de BLSE est parmi les plus élevés du monde. Les types SHV et TEM sont fréquents, CTX-M est endémique et largement dominant. Selon de récentes enquêtes multi-continentales, les Klebsiella isolés de l'Amérique latine ont la prévalence la plus élevée de BLSE (45,4 %). Chez E. coli, elle est de 8,5 % et chez P. mirabilis, 22,4 % (Données programme SENTRY) (Winokur et al. 2001). En 15 ans, la diffusion mondiale des BLSE de type CTX-M chez les entérobactéries a explosé de façon extrêmement rapide, d'où le terme de «pandémie CTX-M» (Rossolini et al. 2008). Les études épidémiologiques récentes rapportent que la situation est endémique dans la plupart des pays d'Europe, d'Asie et d'Amérique du Sud avec de forts taux de prévalence de CTX-M parmi les souches productrices de BLSE : E. coli (de 30 à 90%) et de K. pneumoniae (de 10 à 60 %) (Livermore et al. 2007 ; Rossolini et al. 2008). A noter que quelques CTX-M sont retrouvées spécifiquement dans certains pays (comme CTX-M-9 et CTX-M-14 en Espagne, CTX-M-1 en Italie ou CTX-M-2 en Amérique du Sud, au Japon et en Palestine) tandis que CTX-M-15 est mondialement distribuée et est responsable chez les humains des cas d'infections nosocomiales et communautaires (Rossolini et al. 2008). 32 Chpaitr 1 : Synthese bibliographique 6-2- Les BLSE de type carbapénèmases Les carbapénémases de type KPC (Klebsiella pneumoniae carbapénémase) ont tout d'abord été décrites sur la côte Est des États Unis en 1996 (Yigit et al. 2001) avant d'être identifiées dans la plupart des états du pays, avec notamment une très forte prévalence dans l'état de New York. Ces souches ont également été décrites en Grèce et en Palestine où elles semblent à l'heure actuelle endémiques (Nordmann et al. 2009). Plusieurs variants ont été décrits à ce jour (KPC-1 à KPC-11), le gène blaKPC-2 est actuellement le plus répandu sur la planète. Si les niveaux de résistance aux carbapénèmes des souches productrices d'enzymes de type KPC sont très variables, l'ertapénème est la molécule ayant la plus faible activité. Les souches productrices de KPC sont très fréquemment des souches multirésistantes et les options thérapeutiques sont très souvent réduites. De ce fait, la mortalité associée aux infections par les bactéries productrices d'enzyme de type KPC est élevée, très souvent supérieure à 50% (Nordmann et al. 2011). Les enzymes de type VIM (Verona Integron encoded Metallo-bêta-lactamase) et IMP (Imipénémase) qui représentent la majorité des carbapénémases de classe B ont été rejointes en 2008 par une nouvelle enzyme appelée NDM-1 (NewDelhi metallo-bêta-lactamase 1).La première métallo â-lactamase de type IMP a été décrite au Japon en 1991 (Ito et al. 1995) Depuis, ce type d'enzyme a diffusé dans le monde entier et les enzymes de type VIM et IMP sont désormais endémiques dans certains pays tels que la Grèce, l'Italie, l'Espagne, Taiwan et le Japon (Nordmann et al. 2011). La plupart des souches productrices de métallo-â-lactamases sont des K. pneumoniae multirésistantes acquises dans les établissements de soins. La mortalité associée à ce type de souches varie de 18 à 67% (Nordmann et al. 2011). La première souche de K. pneumoniae productrice d'OXA-48 a été isolée en Turquie en 2003 (Poirel et al. 2004). Depuis, les bactéries productrices d'oxacillinases, notamment OXA-48, ont très largement émergé dans tous les pays du pourtour méditerranéen et en Afrique (Nordmann et al. 2011). Plus récemment, des souches produisant une oxacillinase similaire, OXA-181, ont été isolées en Inde ou chez des patients d'origine indienne (Castanheira et al. 2011). OXA-48 est principalement retrouvée chez K. pneumoniae et E. coli, même si d'autres espèces d'entérobactéries peuvent produire ce type d'enzyme. Les carbapénémases de classe D hydrolysent plus faiblement les carbapénèmes et les céphalosporines de 3ème génération, ce 33 Chpaitr 1 : Synthese bibliographique qui sous-estime probablement leur fréquence de diffusion chez les entérobactéries du monde entier. Toutefois, le niveau de résistance aux carbapénèmes est plus élevé lorsqu'elles sont associées à une BLSE et à un défaut de perméabilité membranaire (Carrer et al. 2010 ; Cuzon et al. 2011). La mortalité associée à ce type d'infection demeure inconnue à ce jour. 7- Facteurs de risque d'acquérir une BLSE Différents facteurs de risque ont été fréquemment associés avec l'acquisition d'une souche productrice de BLSE. Le premier facteur concerne l'utilisation accrue des antibiotiques de type céphalosporines de 3ème génération quelques années avant l'apparition des premières BLSE, et par conséquent, la mise en évidence d'un lien de causalité entre cette utilisation et l'émergence des BLSE (Sirot. 1989). En d'autres termes, les antibiotiques exercent une pression de sélection non-négligeable (Jacobson et al. 1995), et cette pression de sélection est d'autant plus marquée que le nombre de patients traités est important et que la durée de l'antibiothérapie est longue (Asensio et al.2000). De plus, on peut constater que la restriction de l'utilisation des antibiotiques a permis la diminution du nombre de BLSE (Chow et al. 1991 ; Follath et al. 1987). Le deuxième facteur de risque concerne la dissémination des souches résistantes et englobe d'une part le problème des « réservoirs » et d'autre part la transmission des germes. De plus, différents facteurs en relation avec l'acquisition de bactéries productrices de BLSE concerne des patients gravement malades, suite à une hospitalisation prolongée et après exposition à des dispositifs invasifs (cathéters veineux, sonde urinaire ou tube endotrachéal). Un séjour de longue durée implique une plus longue exposition au risque d'acquérir une bactérie multi-résistante comme E. coli et Klebsiella (Kassis-Chikhani et al. 2004 ; Wiener et al. 1999), ce qui signifie une augmentation du risque pour le patient d'être colonisé (Goldsteinet al. 1995). D'autres facteurs de risque sont la malnutrition, l'hémodialyse, la nutrition parentérale totale, l'admission en réanimation ou l'hospitalisation préalable (Lautenbach et al. 2001). |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||