à??°valuation du diagnostic par PCR directe et PCR-élisa sur les ITS des trypanosomes pathogà¨nes du bétail( Télécharger le fichier original )par Bachir SOULEY KOUATO Université Cheikh Anta Diop de Dakar - Doctorat dà¢â¬â¢état en médecine vétérinaire 2005 |

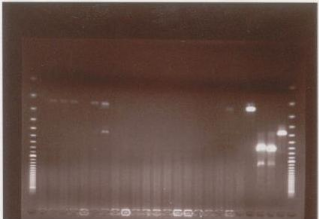
2. Discussion2-1. Evaluation comparative PCR classique / PCR-ITS 2-1-1. Sensibilité Le calcul du Khi 2 pour le total des échantillons positifs en PCR-ITS par comparaison au total des cas positifs détectés en PCR classique (tableau VII, page 84), montre qu'il y a bel et bien une différence entre ces deux techniques et la différence est significative au risque d'erreur 1%. Il ressort donc, au vu de ces résultats que les amorces monospécifiques sont globalement plus sensibles que le couple d'amorces Tryp 4R et Tryp 4S. Une des hypothèses que l'on peut avancer, est la faible répétition des séquences ITS1 dans le génome parasitaire (environ 200 exemplaires) relativement au nombre élevé de copies (un a plusieurs millier) de séquences d'ADN satellite. Par contre, selon les travaux de Desquesnes et al. (2001), la sensibilité de la PCR utilisant les amorces Tryp 4R et Tryp 4S était très voisine de celle obtenue avec les amorces monospécifiques en conditions comparables, mais seulement sur un nombre limité d'échantillons de terrain. Nous avons d'autre part observé que la sensibilité du diagnostic par PCR-ITS sur les buffy coats de bovins s'est avérée inférieure à celle observée avec les techniques parasitologiques (Tableau IV, page 82). Toutefois, les amorces polyspécifiques offrent une sensibilité qui est relative à l'espèce de trypanosome détectée. Ainsi pour les infections de type Duttonella (T. vivax), des meilleurs résultats ont été obtenus avec les amorces polyspécifiques. En effet, les cas positifs à T. vivax représentent presque les 3 / 4 des signaux positifs détectés avec les amorces Tryp 4R et Tryp 4S. Avec les amorces monospécifiques TVW 1&2, la proportion des signaux positifs à T. vivax ne représentent que moins du 1 / 3 des échantillons positifs. Cette sensibilité des amorces polyspécifiques est d'autant plus élevée pour T. vivax que le test de Khi 2 montre des différences significatives entre la détection de cette espèce par la PCR classique et par la PCR-ITS (tableau VII, page 84). Cependant, sur les organes de mouches analysés, le nombre de signaux positifs à T. vivax avec les amorces TVW 1&2 n'est que légèrement inférieur à celui obtenu avec les amorces polyspécifiques. Cette différence est du point de vu statistique non significative (tableau VI, page 83). Contrairement aux amorces Tryp 4R et Tryp 4S, la sensibilité des amorces polyspécifiques Kin 1 et Kin 2 pour la détection de T. vivax est assez faible, car leurs séquences ne présentent que 80 % d'homologie avec celles du parasite (Desquesnes et al., 2001). Pour les infections à T. congolense type savane, les amorces Tryp 4R et Tryp 4S ont donné approximativement le 1 / 4 des résultats obtenus avec le couple d'amorces TCS 1&2 (23 signaux positifs en PCR-ITS contre 99 en PCR classique). Les différences très significatives données par le test de Khi 2 (tableau VII, page 84) traduisent une faiblesse notoire des amorces Tryp 4R et Tryp 4S dans la détection de T. congolense type savane. La technique parasitologique reste donc plus sensible que la PCR-ITS, car la méthode de buffy coat totalise 81 cas d'infections à T. congolense type savane sur 308 prélèvements sanguins de bovins. Toutefois, avec les amorces Kin 1 & 2, la détection de T. congolense type savane est assez bonne. Il faudrait donc envisager, que les amorces Tryp 4R et Tryp 4S ne présentent pas 100 % d`homologie avec toutes les souches de T. congolense type savane. Il y aurait ainsi une différence entre les séquences des souches de T. congolense type savane ayant servi au dessin de ces amorces avec celles des souches présentes sur le terrain. Les résultats des analyses en PCR classique et en PCR-ITS, nous amènent à faire le constat selon lequel, un nombre assez important d'échantillons (21 buffy coats de bovins) positifs à T. vivax en PCR-ITS ont donné des signaux positifs à T. congolense type savane avec les amorces TCS 1et 2. En somme, les amorces Tryp 4R et Tryp 4S présentent un avantage comparatif en gain de sensibilité pour la détection des infections à T. vivax, par contre, la sensibilité relative à la détection de T. congolense type savane de ce couple d'amorces est largement en dessous de celle donnée par les amorces classiques TCS 1& 2. L'interprétation des résultats concernant T. congolense type forêt et T. brucei semble être réservée du fait des faibles taux de détection des infections à ces espèces tant en PCR classique qu'en PCR-ITS. Cela peut s'expliquer par la faible prévalence de ces parasites dans les zones de prélèvements des échantillons. Dans ces zones de suivi du bétail, comme partout ailleurs la présence de T. brucei et de T. congolense type forêt chez les bovins est rarement signalée. Toutefois, la sensibilité des amorces spécifiques de T. congolense type forêt (TCF 1&2) est au regard des résultats obtenus 5 fois supérieure à celle obtenue avec les amorces polyspécifiques. Les taux de détection de T. brucei avec les amorces classiques NRP 1&2 avoisinent légèrement ceux obtenus avec les amorces Tryp 4R et Tryp 4S. Enfin, Les résultats de la PCR classique obtenus dans la présente étude sont comparables et concordent avec plusieurs autres travaux, notamment ceux de Solano et al. (1999). 2-1-2. Spécificité L'un des objectifs primordiaux de l'utilisation des amorces polyspécifiques est de pouvoir distinguer et identifier simultanément les principales espèces de trypanosomes pathogènes du bétail. En effet, les produits d'amplification de l'ADN de ces parasites peuvent être aisément distingués sur un gel d'agarose à 2 % et la taille de ces produits s'est révélée constante et est en accord avec les tailles attendues (Desquesnes et al., 2001). Pour T. vivax, la taille en paires de bases des produits PCR se situe entre 240 et 250 pb, pour les espèces du sous genre Trypanozoon (T. brucei) elle est d'environ 480 pb et enfin pour T. congolense type savane et T. congolense type forêt, les tailles sont respectivement de 680 et 720 pb. Ces tailles sont en dessous de celles observées avec le couple d'amorces Kin 1 et Kin 2 (Desquesnes et al., 2001). Cependant, avec le couple d'amorces Tryp 4R et Tryp 4S on note souvent l'existence de quelques produits non spécifiques dont la taille se situe aux environs de 500 pb, les résultats des PCR avec l'ADN de T. theileri infirment la suspicion selon laquelle ces produits émaneraient d'une infection par cette espèce. Mais la présence de ces produits non spécifiques ne gène aucunement pas la lecture des résultats. Les oligonucléotides classiques offrent une spécificité totale, car aucune réaction croisée entre les trypanosomes n'a été signalée et en plus l'apparition des signaux non spécifiques est relativement moins fréquente. 250 pb 480 pb 620 pb 680 pb MP 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 MP
Figure 23a : Photo d'un gel d'agarose MP : Marqueur de poids, 21 : Témoin positif à T. vivax, 22 : Témoin positif à T. c. s 23 : Témoin positif à T c f, 24 : Témoin positif à T. brucei. 2, 3, 4, 6 et 7 : échantillons positifs à T. vivax 250 pb 480 pb 620 pb 680 pb MP 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 MP
Figure 23b : Photo d'un gel d'agarose MP : Marqueur de poids, 21 : Témoin positif à T. vivax, 22 : Témoin positif à T. c. s 23 : Témoin positif à T c f, 24 : Témoin positif à T. brucei. Entourés bleu : produits non spécifiques. 2-2. Spécificité et sensibilité des sondes nucléiques pour la PCR-ELISA Il est d'abord utile de rappeler que l'une des difficultés de l'application de la PCR-ELISA est indéniablement la recherche des sondes spécifiques aux ITS1 des trypanosomes. Le dessin de ces sondes implique au préalable la connaissance de la séquence du fragment cible. L'obtention des sondes spécifiques aux ITS1 a été ainsi l'aboutissement d'un long processus allant du séquençage des produits d'amplification (avec des amorces polyspécifiques) à l'alignement inter espèces des séquences ITS1 identifiées. 2-2-1. Spécificité Les résultats de l'évaluation des sondes sur des ADN purifiés ont mis en évidence surtout la spécificité complète des oligonucléotides choisis pour le test ELISA. Deux caractéristiques illustrent la stabilité de l'hybridation de ces sondes avec leurs séquences cibles, il s'agit du pourcentage moyen (pour les 5 sondes retenues) de bases GC qui est de 64,48 % (> 60 %) et de la taille moyenne des sondes de l'ordre de 21 pb (> 20 et < 30 pb). Nos résultats sur la spécificité des sondes sont similaires à ceux obtenus avec des sondes dessinées pour la détection des ADN satellites de T. congolense type savane (Sow, 2004), mais aussi avec les résultats des tests de spécificité des sondes pour la détection de T. brucei et de T. vivax (Masaké et al., 2002). Toutefois, dans le cas de notre travail, la spécificité totale des sondes n'a été observée qu'après optimisation de certains paramètres comme la température d'hybridation. Ainsi, pour des températures d'hybridation inférieures à 56°C, des réactions croisées sont observées entre les sondes dessinées pour les trypanosomes de l'espèce T. congolense. Ces hybridations non spécifiques peuvent probablement s'expliquer par le fait qu'en dessous de la température d'hybridation, une sonde a la capacité de s'hybrider avec une autre séquence ITS1 présente dans le milieu réactionnel. Cette hypothèse paraît certainement insuffisante pour justifier à elle seule ce phénomène d'hybridation non spécifique. La vérification de l'homologie entre les sondes d'une espèce donnée et les séquences ITS1 d'une autre espèce s'avère intéressante. Pour éviter ces cas d'hybridations non spécifiques, l'homologie entre un oligonucléotide donné et les séquences des ITS1 des autres espèces doit normalement être inférieure à 80 %.
2-2-2. Sensibilité La sensibilité des sondes à détecter les produits d'amplification des échantillons de terrain est contrairement à toute attente en dessous de celle de la PCR directe, car l'objectif principal de la mise au point et de l'application de la PCR-ELISA est l'amélioration de la sensibilité du test d'identification des trypanosomes par PCR. L'évaluation de la sensibilité des sondes étant réalisée avec des échantillons dont le statut est connu par la révélation sur gel d'agarose, la détection de ces produits PCR par la technique ELISA devrait être théoriquement de 100 %. Par ailleurs, l'on note une différence de sensibilité des oligonucléotides selon l'espèce de parasite à révéler. L'hypothèse la plus plausible qui expliquerait cette sensibilité relative au taxon de trypanosome s'appuie sur un des principes de la PCR-ELISA, à savoir le marquage des produits PCR. En effet, pour la PCR-ELISA, les produits d'amplification sont au préalable marqués avec la digoxigénine (sous forme de digoxigénine-11-2'-dUTP). La digoxigénine est incorporée aux dNTP dans un mélange connu sous le nom de dig-labeling mix qui contient outre les dATP, dCTP et dGTP, les dTTP associés aux dig-dUTP dans une proportion de 19 / 1 (respectivement de dTTP et dig-dUTP), soit un mélange à 5 % de dig-dUTP. Ces derniers vont par le principe de la complémentarité des bases se lier aux bases AT de la séquence ITS1 cible. Ce sont donc, ces complexes qui vont être révélés par le conjugué anti-dig-pod après l'ajout du substrat chromogène. Il faut en plus, notifier que le nombre de bases AT est variable selon la séquence ITS1. Par exemple pour T. vivax dont le produit PCR est le plus court (240 à 250 pb), le nombre de bases AT calculé à l'aide du logiciel FREQSQ est en moyenne égal à 47, ce qui correspond à environ deux Uridine pouvant se fixer. Ceci explique fort probablement la faible sensibilité de la détection des échantillons positifs à cette espèce de trypanosome par ELISA. Par contre, pour T. congolense type savane et T. brucei, on a respectivement 8 (160 bases AT) et 7 (153 bases AT) dig-dUTP pouvant être révélés, d'où leur sensibilité de détection relativement grande par rapport à celle de T. vivax. 2-3. Etude de coût2(*) Pour la réalisation de la PCR-ELISA, il faut un ensemble de réactifs qui serviront d'une part à l'amplification des échantillons et d'autre part à la révélation ELISA. Le coût et la quantité de réactifs seront en premier lieu évalués en fonction du nombre de plaques ELISA utilisées, ce qui conduira au calcul du coût des réactifs par plaque. La plaque ELISA (Maxisorp®) pouvant permettre l'analyse de 45 échantillons (avec un blanc, un négatif et un positif), le coût par plaque ELISA sera divisé par 45 pour obtenir le coût d'analyse d'un échantillon pour une valence (c'est-à-dire la révélation d'une espèce de trypanosome). Avec les amorces Tryp 4R et Tryp 4S, 4 valences sont en jeu, à savoir T. vivax, T. congolense type savane, T. congolense type forêt et T. brucei, pour avoir ainsi le coût d'un diagnostic par PCR-ELISA sur les ITS, on multiplie par 4 le coût d'analyse d'un échantillon sanguin de bovin. Pour la PCR directe, on estime d'abord les prix individuels par unité de consommable à partir des prix unitaires par conditionnement. Puis, on calcule la quantité ou le volume de réactif utilisé pour un gel d'agarose à 2 % sur lequel on peut avoir 39 échantillons amplifiés (avec 2 marqueurs de poids par portion de gel, 4 témoins positifs également par portion de gel et 1 témoins négatif pour le gel entier). Le coût par gel d'agarose d'un réactif donné est obtenu en multipliant son prix individuel / unité par la quantité ou le volume utilisé / gel. Le coût total par gel d'agarose divisé par 39 donne ainsi le prix approximatif de l'analyse d'un échantillon par PCR-ITS directe. Cette technique permettant d'identifier simultanément les 4 espèces de trypanosomes, le coût d'analyse d'un échantillon (buffy coat de bovin) équivaut au coût d'un diagnostic pour 4 valences, par contre pour la PCR classique ce coût est multiplié par 4 comme pour la PCR-ELISA. Certains réactifs dont le prix de revient est très négligeable, n'ont pas été pris en compte dans cette estimation du coût, c'est le cas par exemple du DMSO, du tampon de charge, du marqueur de poids (SL 100), du TBE et du tampon d'hybridation. Ceux-ci sont reconstitués à partir du mélange d'autres réactifs disponibles au laboratoire. Le tampon utilisé en PCR est vendu avec la Taq polymérase (Quiagen®). Enfin, le coût à l'achat des produits comporte également le prix du transport qui représente 25 % du coût du consommable. Ainsi, l'analyse d'un échantillon par PCR-ELISA avec les amorces Tryp 4R et Tryp 4S coûte approximativement 0,45 € soit environ 295,18 Fcfa pour une valence. Le diagnostic chez les bovins avec 4 valences coûterait 1,8 € soit 1180,72 Fcfa. Il en résulte que ce coût est relativement moins onéreux, car dans les mêmes conditions le coût de l'analyse d'un échantillon par PCR-ELISA pour la détection de l'ADN de T. congolense type savane (Sow, 2004) a été estimé à 1993, 29 Fcfa, d'où une réduction de 812,57 Fcfa permise par la PCR-ELISA sur les ITS. Mais cette réduction du coût de la PCR-ELISA, se justifie entre autre par le fait que pour le besoin de notre étude, les réactifs sont achetés séparément. Par contre, dans le cas des travaux de Sow (2004), deux Kits PCR-ELISA (Dig-labeling et Dig-detection 5' Pack) qui sont des produits très chers ont été achetés. Toute chose égale par ailleurs, la PCR-ELISA sur les ITS des trypanosomes offre un avantage du point de vue coût par rapport à la PCR-ELISA pour la détection des ADN satellites, car à la différence de cette dernière, la PCR-ITS permet l'identification simultanée des principaux trypanosomes pathogènes du bétail. L'appréciation du tableau VIII, permet de déduire que le coût de la PCR-ELISA peut être davantage réduit, en diminuant la quantité utilisée de certains consommables comme les tubes eppendorf et les embouts jetables. En effet, si l'on prend l'exemple des tubes eppendorf pour une plaque, ils reviennent plus chers que l'ensemble des six premiers réactifs (la streptavidine, le conjugué anti-dig-pod, le dig-labeling mix, les sondes, le substrat chromogène et les amorces). Par conséquent, pour la révélation ELISA, avec l'utilisation des mêmes tubes ayant servi pour l'amplification et avec une réduction de moitié des embouts jetables, on aboutirait à un coût de 15,33 € par plaque soit donc 0,34 € (223 Fcfa) pour un échantillon analysé. Le coût global du diagnostic chez les bovins avec les 4 valences serait de 1,36 € soit environ 892 Fcfa. Tableau VIII : Estimation du coût de l'analyse en PCR-ELISA pour une plaque
Pour la PCR directe avec le couple d'amorces polyspécifiques (Tryp 4R et Tryp 4S), le coût de l'analyse d'un échantillon est estimé à 0,72 € soit 472,30 Fcfa. Le coût de la PCR monospécifique a été estimé à 0,8 € (Kasbari, 2001). Pour un diagnostic chez les bovins avec 4 valences, la PCR classique serait plus de 4 fois plus coûteuse que la PCR-ITS. Par ailleurs, la PCR-ELISA sur les ITS est 2,5 fois plus chère que la PCR-ITS directe. De ce fait, pour un diagnostic de routine, la PCR-ITS directe constitue une alternative qui doit être privilégiée par rapport à la PCR-ELISA. Tableau IX : Estimation du coût de l'analyse d'un échantillon en PCR-ITS
2-4. Avantages de la PCR directe et de la PCR-ELISA sur les ITS L'intérêt premier des amorces polyspécifiques est sans aucun doute leur capacité à détecter et à identifier simultanément les principales espèces de trypanosomes. De cet avantage découlent d'autres arguments qui militent en faveur de ce nouveau moyen de diagnostic. Ainsi, la réduction du nombre de réactions limite considérablement les cas de contamination, surtout dans les laboratoires moins ou modestement équipés. Cette catégorie de laboratoires est malheureusement caractéristique des pays sous développés où la trypanosomose animale sévit de façon endémique. Du fait de la diversité des espèces de trypanosomes, pour réaliser un diagnostic final avec la PCR monospécifique, il est nécessaire d'effectuer autant de réactions que de nombre d'espèce à identifier. Au demeurant, le coût de la PCR reste onéreux et limite son usage à des fins de diagnostic de routine. La mise au point d'une PCR unique par l'amplification des ITS1 présente alors un avantage tant du point de vue pratique qu'économique. La PCR-ITS favorise entre autres l'analyse d'un plus grand nombre d'échantillons, à moindre coût et en un temps réduit. En outre, les amorces polyspécifiques tiennent une importance de choix dans la connaissance épidémiologique des trypanosomoses animales en permettant d'une part la détection par exemple chez les insectes vecteurs (principalement les glossines) d'autres espèces de trypanosomes autres que les plus fréquemment rencontrées, d'autre part ces amorces constituent des outils les mieux adaptés à la détection des cas d'infections mixtes. Quant à la PCR-ELISA sur les ITS, elle offre les mêmes avantages que la PCR-ITS directe. La particularité de la PCR-ELISA est qu'elle associe deux méthodes de diagnostic à savoir la PCR et la technique ELISA. Elle constitue de ce fait un excellent moyen pour l'amélioration de la sensibilité et de la spécificité des tests de détection des trypanosomes. Cette performance de la PCR-ELISA est illustrée par des travaux effectués sur les ADN satellites (Kosum et al., 2002 ; Masaké et al., 2002 ; Sow, 2004). En outre, cette technique rend la lecture des résultats plus objective et plus aisée grâce à l'utilisation du spectrophotomètre, comparativement à la révélation sur gel d'agarose où l'apparition des bandes est parfois moins nette voire illisible. La détection des produits d'amplification par ELISA est une technique facile à mettre en oeuvre dans les laboratoires où l'on pratique conjointement la PCR et l'ELISA, elle ne nécessite donc aucun investissement supplémentaire. Contrairement à la révélation sur gel d'agarose, dans la PCR-ELISA aucun réactif cancérigène comme le BET n'est utilisé. * 2 Il s'agit ici d'une estimation à minima du coût de la PCR directe et de la PCR-ELISA sur les ITS, sont donc exclues de cette estimation les différentes charges fixes de la PCR. L'objectif de cette étude de coût est d'avoir un aperçu sur une diminution attendue du coût de la PCR -ITS par rapport à celui de la PCR amplifiant les ADN satellites. |
|