CHAPITRE I. INTRODUCTION
1.1. Aperçu général
Les insectes constituent un groupe important de la
biodiversité du règne animal et jouent un rôle
écologique remarquable comme bio-indicateurs de l'état des
écosystèmes (Oudjiane et al. 2014). Ils interagissent
à de multiples niveaux au sein des écosystèmes. Ils sont
alors à l'origine de différents services (Loreau, 2000 ; Lavelle
et al., 2006). D'après Wratten et al. (2007), les
milieux naturels et les agrosystèmes renferment souvent une faune
entomologique typique et riche en espèces.
Plusieurs auteurs tels qu'Andersen et al. (2002),
Hites et al. (2004), Rohr et al. (2006) cité par
Groc(2007) montrent dans leurs études que la reconnaissance des insectes
dans les processus touchant les écosystèmes et comme composants
majeurs de la biodiversité a provoqué progressivement une
augmentation significative de leur représentation. Cependant, leur
incorporation dans les études en particulier pour les
écosystèmes terrestres est plutôt considérée
comme onéreuse (Andersen et al. .2002 ; New, 1996 ; Oliver
& Beattie, 1996 ; Schnell et al., 2003 cité par
Groc,2007).
Par ailleurs, la pollinisation est un facteur clé dans
la reproduction sexuée de nombreuses espèces
végétales. La symbiose entre les insectes et les plantes permet
le maintien de la biodiversité. Il se fait grâce à la
pollinisation croisée et la pérennisation de certains
écosystèmes. Cette activité de pollinisation par les
insectes a une contribution importante dans l'amélioration de la
qualité et de la quantité des fruits et de légumes
(Vaissière, 2005).
Ainsi, la connaissance des insectes pollinisateurs des
différentes plantes cultivées est très importante. La
protection des populations d'insectes pollinisateurs devient alors plus que
nécessaire pour préserver la sécurité alimentaire
et l'état des écosystèmes.
L'Indice Planète Vivante (2020) montre que la
population animale mondiale a diminué de 68 % en moyenne en moins d'un
siècle. Le rapport Global Risks (2020) du Forum Economique Mondial
montre que les cinq défis les plus urgents auxquels l'Afrique sera
confrontée au cours de la prochaine décennie sont tous
liés à l'environnement. Ils incluent la perte de
biodiversité (WWF, 2020). D'après Peter et al. (2012)
à peu près 1,3 million d'espèces des insectes sont
décrites alors qu'ils constituent 55 % de la biodiversité animale
(Mawdsley and Stork, 1995). Leur biomasse est estimée 300 fois
supérieure à celle des autres classes du règne animal.
Selon Wilson (2008), les insectes sont décrits comme « les petites
choses qui font tourner le monde».
2
En Afrique, le déclin des espèces insectes varie
selon les espèces et plusieurs chercheurs s'accordent à dire que
les causes sont presque toutes d'origine anthropique (MTES, 2019).
1.2. La problématique
Bien que la recherche sur la diversité entomologique en
République Démocratique du Congo (RDC) ait connu un essor
significatif depuis deux décennies (Lukoki, 2021), la connaissance de
l'entomofaune demeure fragmentaire et inégalement répartie sur le
territoire national (Milau et al., 2020). À ce jour, un
déséquilibre géographique subsiste : si l'ouest du pays,
notamment Kinshasa et le Kongo-Central, ainsi que la province de la Tshopo ont
bénéficié de nombreuses investigations (Kapiamba, 1980 ;
Badjedjea, 2006 ; Desclée, 2017 ; Bukaka et al., 2020 ; Lukoki et al.,
2021), l'Est du pays, malgré les travaux récents de Rubabura et
al. (2019, 2020, 2021), reste encore largement sous-exploré.
À cette disparité géographique s'ajoute
un biais taxonomique majeur. La majorité des études se sont
focalisées sur des groupes spécifiques tels que les moustiques,
les papillons, les mouches des fruits ou les abeilles, délaissant
d'autres taxons essentiels comme les coléoptères. Pourtant, selon
Biot, (2011) ce manque de données est d'autant plus préoccupant
que les populations d'insectes subissent un déclin mondial sans
précédent sous l'effet de pressions anthropiques multiples.
Dans la province du Sud-Kivu, et plus spécifiquement
dans la région de Lwiro, le déficit de connaissances est
frappant. Si les travaux de Paul et al. (2015) sur la rivière Kalango
ont révélé une abondance exceptionnelle d'insectes dans le
groupement de Bugorhe, leur étude s'est strictement limitée
à la faune aquatique. Or, les insectes pollinisateurs, véritables
piliers de la stabilité des écosystèmes et de la
sécurité alimentaire par leur rôle dans
l'amélioration de la production agricole (Vaissière, 2015), n'ont
fait l'objet d'aucune recherche systématique dans cette
région.
1.3. Questions de recherche
Quand à cette recherche, il nous a été
d'une grande importance d'orienter les questions de la manière suivante
:
· Quels sont les insectes pollinisateurs rencontrés
à Lwiro ?
· Quelle est la composition de la biodiversité des
insectes pollinisateurs de Lwiro ?
· Existe-t-il des biotopes qui soient plus
diversifiés en insectes pollinisateurs que les autres à Lwiro
?
3
1.4. Hypothèses
De ce fait, 3 hypothèses ont été
conçues de la manière suivante ;
· La région de Lwiro serait plus
diversifiée en termes des insectes pollinisateurs qui seraient
composés des hyménoptères, des lépidoptères,
des coléoptères et des diptères,
· Au sein de la population des insectes pollinisateurs
à Lwiro, certains groupes d'insectes seraient plus abondants que les
autres dans les différents les biotopes de Lwiro,
· Certains biotopes de Lwiro seraient plus
diversifiés en insectes pollinisateurs que les autres.
1.5. Objectifs de la recherche 1.5.1. Objectif
principal
L'objectif principal de cette étude est de
caractériser la biodiversité des insectes pollinisateurs dans la
région de Lwiro
1.5.2. Objectifs Spécifiques
? Faire la prospection des biotopes et la captures des
spécimens d'insectes pollinisateurs ? Déterminer les abondances
relatives, des insectes pollinisateurs de la région de Lwiro ainsi que
la richesse spécifique de cette région en insectes
pollinisateurs
? Comparer les différents biotopes (les
jachères, les champs des cultures et milieu naturel) par le biais des
indices de biodiversité tels que l'indice de Shannon-Weaver, l'indice de
Piélou, l'indice de Simpson et l'indice de Sorensen
1.6. Choix du sujet
Le thème a été orienté sur les
insectes pollinisateurs puisqu'ils jouent plusieurs rôles dans la nature.
En effet, ils sont des bio-indicateurs des états des
écosystèmes. Plus nombreux et les plus diversifiés du
règne animal, ils interviennent dans la pollinisation chez les
végétaux ainsi que dans la chaîne tropique. Certaines
espèces sont menacés d'extinction selon la liste rouge de l'UICN
comme le cas des plusieurs espèces d'abeilles sauvages.
4
1.7. Intérêt du sujet
L'intérêt de ce travail a un triple aspect :
1.7.1. Intérêt socio-économique
L'inventaire des insectes pollinisateurs permet leur
conservation puisqu'on ne conserve pas ce qu'on ne connaît pas. Les
insectes pollinisateurs garantissent la sécurité alimentaire par
leur fonction d'améliorer la productivité chez les
végétaux.
1.7.2. Intérêt scientifique
Ce travail met en exergue la richesse spécifique des
insectes pollinisateurs de Lwiro et la manière dont ils interagissent
avec leur environnement. Le taxon des insectes est le plus diversifié du
règne animal.
1.7.3. Intérêt personnel
Personnellement, cette étude va me permettre
d'améliorer mes capacités en zoologie animale
précisément en entomologie
1.8. Période d'étude
Notre période d'étude va d'Octobre 2022 jusqu'en
décembre 2022.
1.9. Subdivision du travail
Ce présent travail est structuré en six
chapitres principaux. Le premier chapitre présente l'introduction
générale, comprenant le contexte de l'étude, la
problématique, les hypothèses, les objectifs ainsi que
l'intérêt scientifique du travail. Le deuxième chapitre est
consacré à la revue de la littérature, dans laquelle sont
développés les fondements théoriques relatifs à la
pollinisation entomophile. Le troisième chapitre décrit le milieu
d'étude et la méthodologie adoptée. Il présente la
région de Lwiro, les caractéristiques des trois biotopes
étudiés (jachère, champ cultivé et milieu naturel),
les techniques d'échantillonnage ainsi que les méthodes d'analyse
des données. Le quatrième chapitre expose les résultats
obtenus, présentés de manière descriptive et analytique
à l'aide de tableaux et figures. Le cinquième chapitre est
consacré à la discussion des résultats, en les confrontant
aux données de la littérature scientifique.
Enfin, le sixième chapitre présente la
conclusion générale du travail ainsi que les recommandations
formulées à l'issue de l'étude.
5
CHAPITRE II. REVUE DE LITTERATURE 2.1. La
pollinisation
2.1.1. Définition de la
pollinisation
La pollinisation est un mode de reproduction des plantes
angiospermes et gymnospermes. Il s'agit du processus de transport d'un grain de
pollen depuis l'étamine (organe mâle) vers les stigmates (organe
femelle). Cela peut se faire soit par autofécondation (concerne une
minorité de plantes telles que les légumineuses ou les
graminées), soit par fécondation croisée
c'est-à-dire que le grain de pollen d'une fleur se dépose sur les
stigmates d'une autre fleur de la même espèce, faisant souvent
intervenir un insecte pollinisateur (Geneves, 1992 ; Pouvreau 2004).
2.1.2. Les modes de pollinisation
Il existe trois modes de pollinisation : l'anémogamie, la
zoogamie et l'hydrogamie
· L'anémogamie : correspond au
phénomène de transport par le vent. Ce dernier va pouvoir
transporter le pollen d'une plante à l'autre. Dans 1/5ème des
cas, la fécondation se fait par cette voie
· L'hydrogamie : correspond au transport du pollen par
l'eau. Ce phénomène reste très marginal et ne concerne que
quelques plantes dont le pollen est de forme très allongée
· La zoogamie : est le transport du pollen par les
animaux. C'est le phénomène le plus courant, il concerne
4/5ème de ce cas de pollinisation. Les insectes contribuent à la
reproduction de 90% des 250000 espèces angiospermes (espèces
à fleurs) recensées dans le monde (Pouvreau, 2004). La
pollinisation par les insectes (dite pollinisation entomophile) résulte
d'un processus de coévolution. Cela se manifeste notamment par une
spécialisation des pièces buccales en fonction du pistil des
plantes à polliniser (Vaissiere et al., 2005). Plus de 1000
espèces de vertébrés participent à la pollinisation
(Colibris, chauves-souris...), auxquels s'ajoutent de très nombreux
insectes. Parmi les insectes, plusieurs ordres sont particulièrement
impliqués, comme les lépidoptères, certains
coléoptères, les diptères (syrphes notamment) et les
hyménoptères (Chagnon, 2008).
6
2.1.3. Importance agro-économique et
écologique de la pollinisation
La pollinisation entomophile est le facteur clé de la
reproduction de la majorité des angiospermes. Ecologiquement, c'est
aussi un élément primordial pour le maintien de la
diversité et de la stabilité des écosystèmes. Elle
permet un meilleur brassage génétique. Celui-ci évite la
dégénérescence. Elle permet une grande diversité et
une résistance accrue des plantes. De ce fait, les insectes
pollinisateurs jouent un rôle très important et
irremplaçable dans l'évolution des plantes sauvages et
cultivées (Vaissiere ,2002).
Par addition près de 75% des plantes angiospermes
dépendent des insectes pollinisateurs pour leur reproduction
sexuée. Les insectes pollinisateurs constituent ainsi un chainon
essentiel dans l'agriculture et la sécurité alimentaire
puisqu'elle forme le vecteur indispensable pour la dissémination de
pollen des espèces végétale (Poureau, 1987 ; Vaissiere
,2002 ; Terzo & Ramont, 2007 ; Fourmier, 2008 ; Praz et al. 2008
cités par Djebli & Nekkeche, 2016).
En fin l'apport des insectes pollinisateurs aux principales
cultures mondiales en 2005 a été évalué à
153 milliards d'euros, ce qui représente 9,5 % de la valeur de la
production alimentaire mondiale (Gallai et al., 2009).
2.2. Les différents ordres des insectes
pollinisateurs
2.2.1. Aperçu général
On distingue quatre ordres d'insectes ayant une réelle
activité sur les fleurs. Le rôle dans la pollinisation des fleurs
est lié à leurs caractéristiques morphologiques, Notamment
leurs pièces buccales. En fonction de la morphologie de celles-ci, les
insectes sont plus ou moins spécialisés dans la pollinisation de
certaines fleurs (Pouvreau, 2004).
2.2.2. Les coléoptères
Bien que les coléoptères constituent 40% des
espèces d'insectes, ils sont les insectes les moins adaptés
à la pollinisation des fleurs, en raison notamment de leurs
pièces buccales courtes et de leur poids important. L'ordre des
Coléoptères regroupe un grand nombre d'espèces xylophages
et phytophages. Les adultes peuvent aussi être floricoles (Brown,
1991)
La pollinisation par les coléoptères est ainsi
assez brutale et peut causer des dommages aux fleurs (stigmates ou pistils
endommagés). Il arrive parfois que ces insectes aient un impact
négatif sur les cultures lorsqu'ils sont trop nombreux comme c'est le
cas des cétoines par exemple (Pouvreau, 2004).
7
Les coléoptères sont caractérisés
par :
· La présence d'élytres, qui sont leurs
ailes antérieures épaisses. Elles protègent effacement
leur abdomen des agressions extérieures.
· Les ailes postérieures, se replient au repos
afin de se détendre.
· Ils disposent de mâchoires puissantes, capables
de broyer de nombreux aliments.
· Ces insectes effectuent une métamorphose
complète. C'est -à-dire qu'ils naissent sous la forme d'un oeuf.
Ils deviennent ensuite une petite larve, puis une nymphe, avant d'atteindre
l'âge adulte. Ces quatre phases de leur vie sont très
régulières chez les coléoptères (Pouvreau,
2004).
2.2.3. Les diptères
Les diptères ont la particularité de ne
posséder qu'une paire d'ailes, la seconde est modifiée et
réduite pour constituer un balancier ou haltère. L'appareil
buccal est généralement de type suceur. On distingue les
Nématocères aux antennes filiformes (moustiques) et les
Brachycères aux antennes courtes et trapues (mouches). Les larves sont
terrestres ou aquatiques, très différentes des imagos dans tous
les cas (120000 espèces dans le monde). Les diptères
possédant un labium court visitent des fleurs aux nectaires accessibles.
Certains ont un labium long qui leur permet d'accéder aux nectars moins
accessibles chez certaines fleurs avec une corolle étroite. Parmi eux
les syrphes sont de bons pollinisateurs (Pouvreau, 2004)
2.2.4. Les lépidoptères
Les Lépidoptères sont un ordre d'insectes
holométaboles dont la forme adulte (ou imago) est communément
appelée papillon et dont la larve est appelée chenille, et la
nymphe chrysalide. Ils se caractérisent à l'état adulte
par trois paires de pattes (comme tous les insectes) et par deux paires d'ailes
recouvertes d'écailles de couleurs très variées selon les
espèces. En outre, Les lépidoptères possèdent une
trompe permettant d'atteindre le nectar de fleurs très étroites,
moins accessible aux autres pollinisateurs. Ils peuvent avoir une
activité nocturne, diurne ou les deux (Pouvreau, 2004)
2.2.5. Les hyménoptères
L'ordre des hyménoptères est celui qui a le plus
d'importance en termes de pollinisation. Le rôle principal est tenu par
la super-famille des Apoïdes (Pouvreau, 2004)). Les Apoïdes
Comprennent les bourdons et les abeilles, soit 20 000 à 30 000
espèces dans le monde. Les abeilles solitaires et sauvages
représentent 85% des Espèces d'Apoïdes. Leur activité
n'est donc pas négligeable et est même plus importante que Celle
des abeilles domestiques (Campbell,
8
1995 ; Pouvreau, 2004). Les abeilles possèdent une
langue (glosse) qui leur permet de récolter le nectar des fleurs.
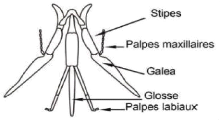
Figure 1. Appareil buccal d'une abeille à
langue longue ( Eardley et al., 2010).
2.3. Relation plantes-pollinisateurs
La relation entre les plantes et les pollinisateurs est l'un
des exemples les plus aboutis de mutualisme dans le monde vivant. Au cours de
l'évolution, les végétaux ont développé des
stratégies d'attraction sophistiquées (signaux visuels, olfactifs
et récompenses trophiques) pour assurer leur reproduction (Chittka et
Schürkens, 2001). On distingue généralement deux
stratégies adaptatives : la généralisation, où la
plante attire un large spectre de visiteurs, et la spécialisation,
où elle fidélise un groupe restreint de pollinisateurs,
augmentant ainsi l'efficacité du transfert de pollen. Dans les cas
d'hyper-spécialisation, comme le complexe figuiers-guêpes
Agaonidae, la survie des deux partenaires devient strictement
interdépendante (Cook et Rasplus, 2003).
2.3.1. Coévolution et adaptations
morphologiques
Les interactions plantes-pollinisateurs sont ancestrales. Des
découvertes de fossiles dans l'ambre suggèrent que ces relations
existaient déjà au Crétacé, il y a environ 100
millions d'années, impliquant initialement des thrips et des
gymnospermes (Peñalver et al., 2012). Cependant, c'est
l'émergence et la domination des angiospermes qui ont
accéléré la diversification des insectes.
Cette coévolution a façonné des
adaptations morphologiques remarquables. Les insectes ont
développé des pièces buccales spécialisées
(proboscis, langues allongées) et des structures de récolte comme
les corbeilles (corbiculae) ou les brosses à pollen sur les pattes
postérieures des Apidae (Michener, 2007). Ces outils permettent une
collecte optimale du nectar et du pollen tout en limitant les dommages aux
fleurs. Toutefois, cet équilibre millénaire est aujourd'hui
menacé. Le déclin rapide des populations d'insectes avec environ
40 % des espèces
9
documentées menacées d'extinction (Potts et al.,
2010) fait peser un risque systémique sur la stabilité des
écosystèmes mondiaux.
2.3.2. Les syndromes de pollinisation
L'ensemble des traits floraux (forme, taille,
accessibilité) constitue ce que les chercheurs appellent les syndromes
de pollinisation (Fenster et al., 2004). Ces traits permettent de
prédire le type de pollinisateurs associés à une plante
:
? Lépidoptères : Attirés par des fleurs
à corolle tubulaire profonde (éperons) nécessitant une
longue trompe.
? Hyménoptères et Diptères :
Préfèrent les inflorescences ouvertes et peu profondes (ex :
Asteraceae) facilitant l'accès au nectar avec une langue courte.
? Coléoptères : Souvent associés à
des fleurs robustes, aplaties et à ovaires protégés,
capables de résister à leurs pièces buccales de type
broyeur (Tautz, 2009).
2.3.3. Signaux visuels et guides
nectarifères
La communication visuelle repose sur une "signalétique"
colorée adaptée au spectre de vision des insectes. Chaque groupe
possède ses préférences : les papillons de jour
privilégient les tons vifs (bleu, rose, violet), tandis que les
papillons de nuit sont attirés par le blanc qui reflète la
luminosité nocturne. Les abeilles, quant à elles, sont sensibles
aux ultra-violets (UV) mais perçoivent le rouge comme du noir (Chittka,
1992).
Pour guider les insectes vers les zones de récolte, de
nombreuses fleurs arborent des guides nectarifères. Ces motifs, souvent
visibles uniquement dans le spectre UV, agissent comme des balises pointant
vers le centre de la fleur. Certaines plantes utilisent même des
changements chromatiques post-fécondation, comme le marronnier dont le
centre passe du jaune au rouge, signalant ainsi aux insectes que la fleur
n'offre plus de récompense (Gori, 1983).
10
CHAPITRE III. MATÉRIELS ET MÉTHODES 3.1.
Milieu d'étude
3.1.1. Localisation et cadre
géographique
La région de Lwiro tire son nom de la rivière
éponyme, un affluent du lac Kivu trouvant sa source dans le marais
Musisi, au sein du Parc National de Kahuzi-Biega (PNKB). Historiquement, cette
zone a été définie pour la première fois comme
entité écologique par Rahm et Dieterlen (1966) lors de leurs
travaux sur les muridés. Située dans le territoire de Kabare,
à environ 50 km au nord de Bukavu, la région englobe les
localités des groupements d'Irhambi et de Bugorhe. Elle est
drainée par un réseau hydrographique dense comprenant, outre la
Lwiro, des affluents tels que la Nyabachiwesa, la Kalengo et la Kamirihembe
(Beatra, 2002).
Géographiquement, elle s'étend entre 28°
45' et 28° 85' de longitude Est et 2° 15' et 2° 30' de latitude
Sud. Son altitude, marquée par un relief de collines et de
vallées, varie entre 1470 m et 2200 m à la lisière du
PNKB, sur une superficie totale de 41 km2. La population,
estimée à environ 152 000 habitants, est majoritairement
constituée d'autochtones (Rapports annuels Bugorhe et Irhambi, 2011).
3.1.2. Climat, sol et
végétation
Le milieu repose sur des sols d'origine volcanique
particulièrement fertiles. Le climat est de type tropical humide,
caractérisé par une alternance saisonnière marquée
: une longue saison des pluies de neuf mois (septembre à mai) et une
saison sèche de trois mois (juin à août). Avec une
température moyenne annuelle de 19,5 °C et une humidité
relative oscillant entre 68 % et 75 %, la région offre des conditions
optimales pour une grande diversité agricole. La
végétation originelle, aujourd'hui fragmentée, est
dominée par une savane herbeuse de montagne parsemée d'arbustes
et de graminées diversifiées (Beatra, 2002).
Sur le plan institutionnel, Lwiro est un pôle
scientifique historique abritant le Centre de Recherche en Sciences Naturelles
(CRSN, ex-IRSAC) et l'Institut National pour l'Étude et la Recherche
Agronomique (INERA) à Mulungu (Balagizi, 2010).
11
3.1.3. Description des sites d'échantillonnage
(Biotopes)
Pour cette étude, trois biotopes d'une superficie d'un
hectare chacun ont été sélectionnés afin de
comparer la diversité des pollinisateurs.
1. Le biotope de jachère
Situé dans le domaine de Muller à Bishibiru
(02° 14' 33,0" S ; 28° 48' 18,3" E), ce site se trouve à une
altitude de 1708 m. La végétation y est spontanée et
diversifiée, dominée par des espèces telles que
Viguiera dentata, Gymnanthemum coloratum, Bidens pilosa, Lantana camara,
Ocimum gratissimum, Pentas lanceolata, Kitaibelia vitifolia et Ipomoea
asarifolia.
2. Le biotope de culture
Ce site est localisé dans la zone dite «
Magnétisme » à 1687 m d'altitude (02° 15' 06,4» S
; 28° 48' 22,8» E). Contrairement au précédent, ce
milieu est anthropisé et caractérisé par une faible
diversité floristique, principalement représentée par les
cultures de haricot (Phaseolus vulgaris) et de maïs (Zea
mays).
3. Le biotope de milieu naturel
Le choix s'est porté sur le marais de Mbayo,
situé à une altitude plus élevée de 2122 m
(02° 15' 31,8" S ; 28° 45' 18,5" E). Ce milieu humide présente
une flore spécifique adaptée, comprenant notamment Carex
oshimensis, Solanum mauritianum, Canna indica, Zantedeschia aethiopica et
Bidens pilosa.
La Figure ci-dessous présente la localisation
cartographique de nos trois biotopes dans le territoire de Kabare, Province du
Sud-Kivu.
L'échantillonnage s'est déroulé sur une
période de deux mois, s'étendant du 10 octobre au 2
décembre 2022, couvrant ainsi une phase de floraison importante dans les
trois biotopes d'étude.
12
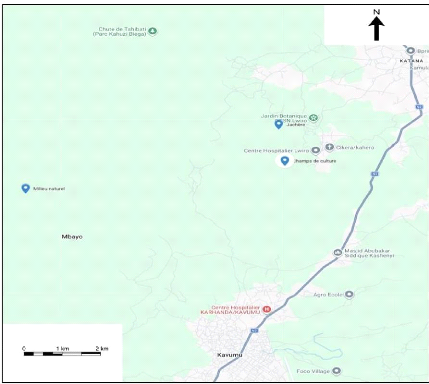
Figure 2. Localisation de sites
d'échantillonnage
Sur la figure 2, les trois points bleus avec un petit trou blanc
représentent nos trois biotopes étudiés
3.2. Méthodologie
3.2.1. Échantillonnage et collecte des
données
L'inventaire de l'entomofaune a été
réalisé par une méthode de capture active (Louis-Michel et
al., 2009). Cette technique consiste à prélever les insectes
directement sur la strate herbacée et arbustive à l'aide d'un
filet fauchoir. Ce choix méthodologique est justifié par le
caractère principalement ailé et la grande mobilité des
insectes pollinisateurs ciblés.
13
Les spécimens capturés ont été
traités selon leur groupe taxonomique :
? Insectes généraux : Conservés dans des
flacons contenant de l'éthanol à 96 % pour une conservation
optimale des caractères morphologiques.
? Lépidoptères : Placés individuellement
dans des enveloppes en papier cristal (papillotes) afin d'éviter la
dégradation de l'écaillage alaire, essentiel pour
l'identification.
3.2.2. Identification taxonomique
L'identification a été réalisée au
Laboratoire d'Entomologie Agricole du Centre de Recherche en Sciences
Naturelles (CRSN/Lwiro). Nous avons adopté l'approche Rapid Biodiversity
Assessment (RBA), qui permet de classer les spécimens par
morpho-espèces afin d'obtenir un relevé de biodiversité
complet, même en l'absence de révisions taxonomiques
récentes pour certains groupes complexes (Krell, 2004).
La détermination systématique s'est appuyée
sur :
? Des clés de détermination
spécialisées (Delvare et Aberlenc, 1989 ; Mike et al., 2004). ?
La comparaison avec les spécimens de référence
conservés dans l'insectarium du CRSN/Lwiro.
Pour l'identification de la flore associée, nous avons
combiné l'utilisation d'outils numériques de reconnaissance (Tela
Botanica, PlantNet) et la consultation des planches de l'Herbarium du CRSN.
3.2.3. Analyse statistique des données
Pour caractériser la structure et la diversité
des peuplements de pollinisateurs, les indices suivants ont été
mobilisés :
3.2.3.1. Abondance relative (Pi)
Elle exprime la proportion d'une espèce par rapport
à l'effectif total et permet de définir la composition de la
biodiversité de chaque biotope (Zaime et Gautier, 1989, cité par
Noudjoud, 2006) :
????.100
Pi=
N
Où ni représente l'effectif de l'espèce i et
N l'effectif total toutes espèces confondues.
14
3.2.3.2. Indice de diversité de Shannon-Weaver
(H')
Cet indice mesure la complexité et la diversité
spécifique du milieu en tenant compte à la fois de la richesse et
de l'équitabilité (Ramade, 2003) :
Où H' est l'indice de Shannon-Waever et Pi est la
fréquence relative de l'espèce i. La valeur maximale de cet
indice (H max) est atteinte lorsque toutes les espèces ont la même
abondance, soit Hmax = ln(S), où S est la richesse spécifique
(Marcon, 2010).
3.2.3.3. Équitabilité de Pielou
(J')
Elle permet d'évaluer la régularité de la
répartition des individus au sein des espèces d'un biotope
(Marcon, 2010).
Avec :
? J' : l'indice de Piélou
? H' : l'indice de Shannon-Weaver
? S : la richesse spécifique d'un point
L'indice J varie de 0 à 1 ; une valeur proche de 1 indique
que les espèces ont des abondances
équilibrées au sein du milieu.
3.2.3.4. Indice de similarité de Sorensen (S ou
IC)
Utilisé pour mesurer la diversité bêta, cet
indice compare la composition faunistique entre deux biotopes (Triplet, 2020)
:
2 A
K = ( 2 A + ?? + ?? ) * 100
Avec:
· A : le nombre d'espèces communes aux deux biotopes
comparés
· B : le nombre d'espèces présentes dans le
biotope B mais absentes dans le biotope C
· C : le nombre d'espèces présentes dans le
biotope C mais absentes dans le biotope B
15
CHAPITRE IV. PRESENTATION DES RÉSULTATS 4.1. Etat
des lieux et Structure de l'entomofaune
Ce premier axe d'analyse dresse le profil taxonomique global
de l'entomofaune pollinisatrice de Lwiro. Il s'agit de caractériser la
structure fondamentale du peuplement afin d'identifier les groupes biologiques
dominants de la région. Cet état des lieux constitue le socle
nécessaire à la compréhension des équilibres
écologiques avant toute analyse comparative des milieux.
4.1.1 Composition taxonomique globale
L'inventaire global réalisé dans la
région de Lwiro a permis de recenser une communauté
composée de 4 ordres, répartis en 10 familles pour un total de 34
espèces, totalisant un effectif de 554 individus (Tableau
1).
Tableau 1. Inventaire taxonomique et abondance
relative de l'entomofaune pollinisatrice dans la région de
Lwiro
Ordres Familles
|
Nombre d'espèces
|
Effectif Abondance relative (%)
|
Hymenoptera
Lepidoptera
Coleoptera Diptera
Total
|
Apidae
|
8
|
110
|
19,86
|
|
5
|
88
|
15,88
|
|
2
|
109
|
19,68
|
|
3
|
17
|
3,07
|
|
1
|
2
|
0,36
|
|
7
|
44
|
7,94
|
|
3
|
118
|
21,30
|
|
1
|
5
|
0,90
|
|
2
|
28
|
5,05
|
|
2
|
33
|
5,96
|
|
34
|
554
|
100,00
|
|
De plus, l'analyse de la structure de ce peuplement,
basée sur les abondances relatives des familles, met en évidence
une répartition inégale des abondances. La communauté est
numériquement dominée par quatre familles majeures qui
représentent, à elles seules, plus de 75 % de
l'échantillon total : les Pieridae (21,30 %), les Apidae (19,86 %), les
Formicidae (19,68 %) et les Vespidae (15,88 %). Ces groupes constituent le
socle de l'entomofaune pollinisatrice locale (Tableau 1).
16
À l'inverse, les autres familles identifiées
présentent une représentativité nettement plus faible. Les
Nymphalidae (5,96 %), les Syrphidae (5,96 %) et les Scarabaeidae (5,05 %)
apparaissent comme des composantes secondaires du système. Les familles
les moins fréquentes, telles que les Lycaenidae, noctuidae contribuent
que de manière marginale à l'abondance globale, chacune
représentant moins de 3 % de l'effectif total (Tableau
1).
Les proportions des ordres de l'entomofaune de Lwiro à
l'échelle globale sont représentées par la figure
ci-dessous :
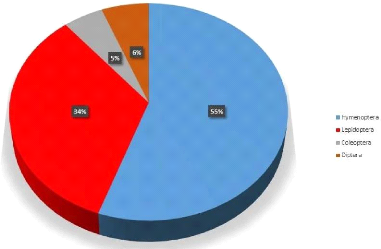
Figure 3. Proportions des ordres de l'entomofaune de
Lwiro
L'analyse du diagramme circulaire révèle une
prédominance marquée des Hyménoptères, qui
représentent à eux seuls 55 % de l'abondance totale du
peuplement. Les Lépidoptères constituent le second pôle
d'importance avec 34 % de l'effectif global. Enfin, les ordres des
Diptères (6 %) et des Coléoptères (5 %) complètent
la structure de la communauté (Figure 1).
.
17
4.1.2. Richesse spécifique et
abondance
L'analyse de la structure de la communauté,
illustrée par la courbe de rang-abondance de whitaker (Figure
4), permet d'appréhender l'organisation écologique du
peuplement au-delà de sa simple composition taxonomique. Cette courbe,
représentée sur une échelle logarithmique, offre une
vision précise du partage des ressources entre les 34 espèces
recensées.
Rang de dominance
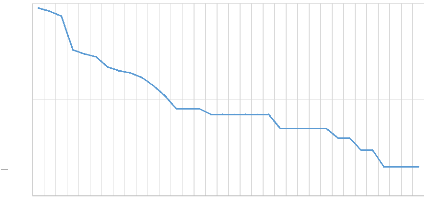
Nombre d'individus (log10)
100
10
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
23 24 25 26 27 28 29 30 31 32 33 34
Figure 4. Structure de la communauté et
distribution des abondances
Le profil graphique met en évidence une structure
typique des écosystèmes tropicaux, caractérisée par
une forte disparité entre les taxons dominants et les taxons satellites.
La partie initiale de la courbe présente une pente abrupte,
révélant la présence de quelques espèces
ultra-dominantes qui concentrent l'essentiel de la biomasse numérique.
Ce phénomène de dominance est particulièrement
marqué pour les trois premiers rangs, correspondant aux espèces
Dorylus niarembensis (Formicidae), Leptidea sinapis
(Pieridae) et Apis mellifera (Apidae).
À l'opposé, la courbe s'étire en une
"longue traîne" horizontale à mesure que l'on progresse vers les
rangs supérieurs (du rang 22 au rang 34). Cette morphologie graphique
témoigne d'une proportion importante d'espèces rares,
représentées par un très faible nombre d'individus,
souvent des singletons. Les rangs des toutes les espèces sont repris en
annexe 9.
18
4.2. Ecologie comparative de biotopes
Pour mieux comprendre comment les insectes pollinisateurs se
répartissent dans le paysage de Lwiro, nous avons comparé la
richesse spécifique au sein de trois biotopes distincts : les
jachères, les champs de cultures et le milieu naturel. Cette approche
permet de mesurer l'attractivité de chaque milieu et d'identifier
précisément ceux qui abritent la plus grande diversité
d'espèces au moment de l'étude (Tableau 2).
Tableau 2. Richesse spécifique (S) par ordre
taxonomique au sein des trois biotopes.
Ordres Jachères (S)
|
Champs des cultures (S)
|
Milieu naturel S)
|
Hymenoptera Lepidoptera Coleoptera Diptera
Total (Richesse S)
|
14
|
3
|
2
|
|
4
|
5
|
|
1
|
0
|
|
1
|
0
|
|
9
|
7
|
|
Les données recueillies montrent une nette
prédominance de la jachère, qui concentre à elle seule 31
espèces, soit la grande majorité de la diversité totale
recensée. Les ordres des Hyménoptères et des
Lépidoptères y sont particulièrement bien
représentés avec respectivement 14 et 13 espèces
identifiées. En revanche, on observe un fléchissement
marqué de la richesse spécifique dans les deux autres milieux,
les champs de cultures ne comptant que 9 espèces et le milieu naturel
seulement 7. Il est également à noter que les Diptères et
les Coléoptères sont totalement absents du milieu naturel dans
nos relevés, tandis que les Hyménoptères restent le seul
groupe présent de manière constante dans l'ensemble des biotopes
étudiés, bien que leur nombre d'espèces diminue de
moitié en dehors des jachères (Tableau 2).
4.3. Analyse de la stabilité et de la
similarité écologique
L'analyse de la stabilité et de la similarité
écologique permet de passer d'un simple inventaire à une
compréhension réelle de la dynamique des milieux. L'objectif est
ici d'évaluer la santé interne de chaque biotope et de mesurer
les liens qui les unissent, afin de déterminer si
l'écosystème de Lwiro forme un ensemble cohérent et
résilient pour les insectes pollinisateurs.
19
4.3.1. Equilibre des milieux
Pour évaluer la structure interne de nos peuplements
d'insectes, il est nécessaire d'aller au-delà de la simple
présence d'espèces et d'analyser la manière dont les
individus se répartissent entre elles. L'utilisation conjointe des
indices de Shannon, de Simpson et de Pielou permet d'appréhender cet
équilibre sous trois angles complémentaires : la
complexité du milieu, la probabilité de dominance d'un groupe
précis et la régularité de la distribution globale. Cette
analyse structurelle est déterminante pour juger de la capacité
d'un biotope à maintenir une biodiversité fonctionnelle et
résiliente
3,13
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1,85
|
|
|
|
|
|
0,95
|
0,91
|
|
|
|
1,3
|
|
|
|
|
|
0,81
|
0,84
|
|
0,67
|
0,67
|
|
|
|
|
|
|
|
JACHÈRES CHAMPS DE CULTURE MILIEU NATUREL
indice de shannon indice de simpson Indice de pielou
Figure 5. Indices de Shannon, de Pielou et de
Simpson dans chaque biotope
L'analyse de la figure 5 montre que la jachère est le
milieu le plus riche et le mieux structuré de notre zone d'étude.
Avec un indice de Shannon de 3,13 et une
équitabilité de Pielou de 0,91, on comprend que
les populations d'insectes y sont réparties de manière
très homogène. L'indice de Simpson vient confirmer ce constat :
il indique une faible dominance, ce qui signifie qu'aucune espèce ne
supplante les autres. La jachère offre donc un environnement stable
où la diversité peut s'exprimer pleinement.
20
À l'inverse, le milieu naturel apparaît comme le
plus fragile. Son indice de Shannon est bas (1,3) et son
équitabilité ne dépasse pas 0,67. Ici,
l'indice de Simpson suggère qu'un petit nombre d'espèces domine
largement le groupe, rendant l'écosystème moins diversifié
et plus instable. Enfin, les champs de cultures présentent un profil
intermédiaire. Bien que leur richesse soit modeste avec un Shannon de
1,85, l'indice de Pielou de 0,84 et l'indice
de Simpson de 0,81 montrent que les insectes présents y
vivent tout de même de façon assez équilibrée, sans
qu'une espèce ne devienne envahissante
4.3.2. Similarité et
connectivité
Après avoir étudié l'équilibre
interne de chaque milieu, il est crucial de comprendre comment ces biotopes
communiquent entre eux. L'indice de Sorensen nous permet ici de mesurer la
parenté faunistique, c'est-à-dire le degré de ressemblance
entre les communautés d'insectes de deux sites différents. En
identifiant la proportion d'espèces partagées, nous pouvons
évaluer la connectivité du paysage : plus l'indice est
élevé, plus les échanges d'espèces sont
facilités, ce qui témoigne d'une continuité
écologique entre les habitats.
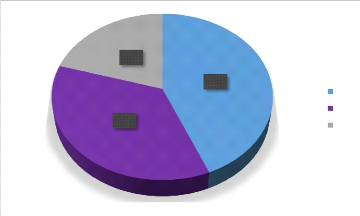
37%
21%
45%
J-C
J-M
C-M
Figure 6. Pourcentage de similarité entre les
biotopes
Les résultats de l'indice de Sorensen mettent en
évidence des niveaux d'affinité très variables selon les
milieux comparés. La similarité la plus forte est observée
entre les jachères et les champs de cultures, avec un taux de 45 %. Ce
chiffre indique que près de la moitié des espèces sont
communes à ces deux habitats, suggérant une circulation fluide
des pollinisateurs entre les zones en repos et les zones cultivées
(Figure 6).
21
À l'opposé, le lien entre les champs de
cultures et le milieu naturel est le plus faible, ne s'élevant
qu'à 21 %. Cette faible similarité montre que ces deux biotopes
abritent des communautés très différentes, avec peu
d'espèces capables de passer de l'un à l'autre. Enfin, la
comparaison entre la jachère et le milieu naturel révèle
une affinité intermédiaire de 37 % (Figure
6).
22
|