Evaluation du potentiel ovarien pour la production in vitro d'ovocytes fécondables dans le plateau de l'Adamaoua( Télécharger le fichier original )par Dawaye Souley monglo Université de Ngaoundere - Docteur en médecine vétérinaire 2014 |

INTRODUCTIONL'élevage bovin au Cameroun occupe une place importante dans le système de production animale nationale. Avec environ 7 millions de têtes, les bovins représentent 10 % du cheptel global des animaux (MINEPIA, 2009). Les bovins élevés au Cameroun sont pour la plupart les zébus (Bos indicus). Les taurins (Bos taurus) représentent à peine 2% de la population bovine totale et sont menacés d'extinction (Lhoste, 1991). A côté de celles-ci, se trouvent également des races exotiques. Toutefois, la productivité de ces races reste faible. Les problèmes génétiques, zootechniques, sanitaires et de reproduction ont été identifiés comme étant les facteurs responsables de la faible productivité (Ebangi et al., 2011). Les bovins locaux camerounais sont élevés selon un mode traditionnel avec peu de programmes d'amélioration génétique. L'insémination artificielle a été introduite pour la première fois en Afrique en 1935 au Kenya, puis s'est généralisée dans toute l'Afrique subsaharienne à la faveur des différents projets d'amélioration (Kouamo et al., 2009). Au Cameroun, elle est encore peu utilisée. Certains éleveurs pratiquent l'insémination sur les races locales avec une semence importée. Mais, cette technique est utilisée de manière anarchique, ce qui entraîne la dispersion et la dilution du génotype local (Bah et al., 2010). La production in vitro d'embryon et le transfert d'embryon sont des techniques de reproduction qui constituent une alternative à l'IA pour l'amélioration génétique (Huang et Rosenwarks, 2012). En effet, ils permettent la préservation du potentiel génétique d'animaux sub-fertiles ou morts (Deuleuze et al., 2009) par la création d'une banque de gènes (Seidel et Seidel, 1989). Les ovaires prélevés sur des animaux après abattage constituent la plus grande source d'ovocytes primaires obtenus à moindre coût pour la production à grande échelle d'embryons bovins suite à la maturation et à la fécondation in vitro (Agrawal et al., 1995). L'étape initiale et primordiale dans la fécondation in vitro est la collecte et la sélection des ovocytes viables capables d'être maturés et fécondés in vitro. A notre connaissance, cette étude n'a jamais été menée au Cameroun. C'est dans ce contexte qu'a été réalisée cette étude avec pour objectif principal l'évaluation du potentiel ovarien des zébus Akou, Bokolo, Djafoun et Goudali pour la production in vitro d'ovocytes fécondables. Il s'agit de façon spécifique de : - caractériser les vaches abattues ainsi que leurs ovaires ; - déterminer la population folliculaire ; - déterminer le rendement et la qualité des ovocytes ; 2 - évaluer les effets des facteurs ovariens et non ovariens sur la population folliculaire, le rendement et la qualité des ovocytes CHAPITRE I : REVUE DE LA LITTÉRATURE A. ANATOMIE ET PHYSIOLOGIE DE L'OVAIRE L'ovaire est un organe qui a quatre fonctions principales à savoir : la multiplication des cellules somatiques (cellules folliculaires) entourant l'ovocyte, la multiplication des ovogonies et la maturation des ovocytes, l'ovulation et la synthèse des hormones stéroïdiennes et peptidiques contrôlant le fonctionnement ovarien mais aussi celui de tous les organes qui interviennent dans la fonction de reproduction de la femelle. I. Anatomie de l'ovaire I.1. Morphologie L'ovaire de la vache (photo n° 1) a la forme d'une amande aplatie latéro-médialement (Cuq et Agba, 1975) avec deux pôles inégaux ; elle peut souvent être sub-ovoïde. La surface de l'ovaire est irrégulière, elle est bosselée par des structures telles que les follicules à différents stades de croissance et le corps jaune. Les dimensions de l'ovaire des bovins rapportées dans la littérature chez Bos indicus sont de 26 à 28 mm pour la longueur, 17 à 18 mm pour la largeur et 13 à 14 mm pour l'épaisseur (Cuq et Agba, 1975). Le poids moyen des ovaires est de 4,42 g pour le zébu Ankolé (Natumyana et al., 2008) et de 10,2 g pour le taurin Swedish Red ( Rajoski, 1960).
3 Photo n ° 1 : Ovaire d'une vache
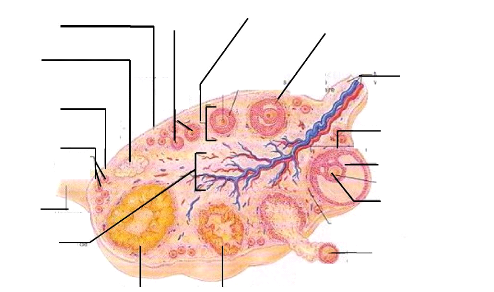
En coupe longitudinale (figure 1), on observe trois parties distinctes de l'ovaire à savoir l'épithélium germinatif qui est la couche externe, l'albuginée constituée de tissus conjonctifs denses et le stroma qui se divise en deux parties : le cortex (couche externe) et la médulla (couche interne). Le cortex est un tissu de soutien formé de tissu conjonctif et de cellules du stroma ; il contient les organites ovariens (follicules, corps jaune). Quant à la médulla, encore appelée zone vasculaire, elle est composée de tissu ovarien nutritionnel et de soutien (Marieb, 1999).
Cortex Follicules primaires Albuginée Follicule secondaire Corpus albican Mésovarium et vaisseaux sanguins Epithélium germinatif Follicule mûr Follicules primordiaux Antrum Ligament propre Ovocyte Medulla Ovocyte expulsé
Figure 1: Structure histologique d'un ovaire Source : Marieb, 1999. Anatomie et Physiologie humaines, p. 1056. 5 I.4. Irrigation et innervation Les ovaires sont irrigués par l'artère ovarique issue de la partie caudale de l'aorte abdominale. La veine ovarique draine l'ovaire du cortex vers la médulla. Elle se jette rapidement dans la veine cave caudale pour la veine ovarique droite, et dans la veine rénale pour la veine ovarique gauche. Les nerfs sont représentés par de nombreux rameaux grêles, anastomosés, constituant le plexus ovarique. II. Physiologie de l'ovaire Au sein de l'ovaire, deux processus de développement étroitement imbriqués, l'ovogenèse et la folliculogenèse, déterminent le nombre et la qualité des ovocytes produits. Ces processus, initiés pendant la vie foetale, se poursuivent pendant toute la vie de la femelle et sont étroitement contrôlés à chacune de leurs étapes par de nombreux facteurs hormonaux et environnementaux (Monniaux et al., 2009). II.1. Ovogenèse L'ovogenèse est la formation des cellules sexuelles femelles, aussi appelées ovules (Marieb, 1999). Elle se déroule dans les ovaires et débute durant la vie embryonnaire (Mauléon, 1961). Elle est d'autant plus complexe qu'elle s'imbrique dans un autre processus de développement, la folliculogenèse, avec laquelle elle entretient des liens étroits (Monniaux et al., 2009). Le processus de l'ovogenèse se déroule en cinq phases et s'échelonne sur plusieurs années. La différentiation sexuelle embryonnaire est établie vers la 6e semaine de la gestation chez la vache. Durant cette période foetale, les cellules germinales primordiales colonisent la crête génitale (phase 1). Elles donnent, après différentiation, naissance aux ovogonies (phase 2) qui sont des cellules diploides (Namdori et al., 2008). Ces ovogonies se multiplient de façon mitotique dans les cordons ovigères jusqu'à leur entrée en prophase méiotique qui marque l'arrêt de leur accroissement numérique (Monniaux et al., 2009). Cette multiplication s'étend du 60e au 170e jour du développement embryonnaire et foetal (Erickson, 1966). Les ovaires peuvent contenir alors jusqu'à 2 millions d'ovogonies (Hanzen et al., 2000) durant la vie foetale. La phase mitotique terminée, ces dernières entament le processus de méiose qui s'interrompt au stade diplotène (dit aussi vésicule germinale) de la prophase I et deviennent ainsi les ovocytes I (phase 3). Chaque ovocyte s'entoure d'une couche de cellules somatiques, de cellules de la granulosa et d'une lame basale pour former des follicules primordiaux (Drion et al., 1996; Monniaux et al., 2009). 6 A la puberté qui intervient à l'âge de 3 à 4 ans chez les zébus (Messine et al., 2003), plusieurs ovocytes I seront activés, un seul sera « choisi » pour poursuivre la première division de la méiose I. Une fois la première division méiotique terminée, l'ovocyte I donne deux cellules haploïdes de volume très inégal. La plus petite des cellules est appelée globule polaire I et la plus grosse, qui contient tout le cytoplasme l'ovocyte II (phase 4). Cet ovocyte va commencer la seconde division de la méiose mais ne la complétera pas. Elle sera bloquée en métaphase II jusqu'à la libération de l'oeuf mature (phase 5). C'est lors de la fécondation ou de l'activation parthénogénétique de l'oeuf que la méiose se termine. En cas de non fécondation, l'ovule dégénère (Marieb, 1999). II.2. Cycle sexuel Un cycle sexuel est l'ensemble d'événements biologiques précis intervenant à trois niveaux: cellulaire, hormonal et comportemental. Il est caractérisé par la croissance et la régression des follicules et du corps jaune (Sartori et Barros, 2011). Sa durée moyenne est de 22,1#177;1,5 jours chez la femelle Zébu (Marichatou, 2010). II.2.1. Evénement cellulaire ovarien Au niveau cellulaire, il y a deux phases conduisant à des formations ovariennes : une phase folliculaire correspondant à la croissance des follicules et qui se termine par l'ovulation ou rupture du follicule et une phase lutéale au cours de laquelle le corps jaune issu de la rupture du follicule croit puis régresse (Stouffer, 2006). II.2.1.1. Folliculogenèse Le follicule est une structure en forme de sac, formée par un amas de cellules mésodermiques, entourant une cellule ou un autre ensemble de cellules. La folliculogenèse commence pendant le développement foetal. En effet, les follicules antraux sont observés dans les ovaires des foetus bovins très tôt à 5 mois de gestation (Santos et al., 2013). C'est un processus continu initié à partir de la réserve de follicules primordiaux jusqu'à l'ovulation ou, cas le plus fréquent, à la dégénérescence (ou atrésie) de plus de 99% des follicules en croissance (Driancourt et al., 2001; Monniaux et al., 2009). L'initiation de la croissance folliculaire se caractérise par l'augmentation du volume de l'ovocyte et l'entrée en prolifération des cellules de la granulosa (Monniaux et al., 2009. Cette phase ne concerne que 10 % du stock folliculaire qui est de 150 000 à 235 000 chez la vache (Hanzen et al., 2000). Elle est caractérisée par des modifications qui concernent à la fois le follicule et l'ovocyte qu'il renferme (Monniaux et al., 2009). 7 II.2.1.1.1. Aspects morphologiques du développement folliculaire La folliculogenèse comprend les stades des follicules primordial, primaire et secondaire, constituant les follicules pré-antraux, puis les stades tertiaire et de De Graaf représentant les follicules antraux (Hanzen et al., 2000; Monniaux et al., 2009).
Il résulte de la transformation du follicule primordial par augmentation du volume de l'ovocyte I, qui est entouré d'une couche de cellules cubiques. Ces cellules synthétisent des glycoprotéines qui formeront la zone pellucide. D'une épaisseur d'environ 10 microns, elle est constituée à 95 % de trois glycoprotéines, organisées en longs filaments interconnectés, appelées ZP1, ZP2, ZP3. La taille du follicules primaire atteint 60 à 80um et celle de l'ovocyte qu'ils contiennent 30 à 40 micromètres (Hanzen et al., 2000).
Figure 3 : Follicules primaires Source : http://mapageweb.umontreal.ca/cabanat/bio2460/Gametogenese.html c) Follicule secondaire Il est constitué d'un ovocyte volumineux, enveloppé d'une zone pellucide complètement différenciée. Celle-ci est entourée de plusieurs couches de cellules cubiques formant la granulosa. La granulosa est limitée à l'extérieur par la membrane de Slavjansky. L'ovocyte atteint à ce stade son volume maximum. Le diamètre du follicule secondaire est compris entre 200 et 400 um et celui de l'ovocyte est d'environ 60 um (Hanzen et al., 2000).
8 Figure 4 : Follicule secondaire Source : http://mapageweb.umontreal.ca/cabanat/bio2460/Gametogenese.html
Il représente la phase terminale du développement folliculaire. Cette phase ne concerne qu'un follicule sur 1000 entrés en croissance. Ce follicule ne peut être observé que pendant le proestrus (Cuq et Agba, 1975). C'est au cours de cette phase que les cavités finissent par confluer pour former l'antrum. Le développement progressif de l'antrum entraîne la séparation des cellules de la granulosa qui se différencient en outre en corona radiata, couche cellulaire entourant directement l'ovocyte et en cellules du cumulus oophorus. La taille du follicule mûr est de 10 à 12 mm chez le zébu (Sartorelli et al., 2005).
Zone granulosa Cumulus oophorus Corona radiata Ovocyte Antre et liquide folliculaires Thèque interne Thèque externe 10 Figure 6: Follicule mûr ou de De Graaf Source : http://mapageweb.umontreal.ca/cabanat/bio2460/Gametogenese.html Tout au long du développement folliculaire, l'ovocyte et les cellules de la granulosa qui l'entourent gardent un contact étroit grâce à l'existence de prolongements cytoplasmiques des cellules de la granulosa qui traversent la zone pellucide et viennent s'apposer contre la membrane plasmique de l'ovocyte. La présence de jonctions communicantes «gap junctions» à ces niveaux de contact est responsable d'un véritable couplage métabolique entre ces deux types cellulaires, permettant des échanges d'ions et de petites molécules de poids moléculaire inférieur à 1 kD (Monniaux et al., 2009). II.2.1.1.2. Dynamique de la croissance folliculaire La population des follicules ovulatoires se renouvelle au cours du cycle oestral par une succession de croissance ou de régression folliculaires successives (Sartori et Barros, 2011 ; Ghinter et al., 2014) appelées « vagues folliculaires ». a) Vagues de croissance folliculaires Une vague folliculaire correspond à la croissance synchrone d'une cohorte de follicules, suivie de la sélection d'un ou de plusieurs follicules qualifiés de follicules dominants et de leur évolution vers l'ovulation, ou de leurs régressions successive quand les conditions endocriniennes sont défavorables (phase lutéale du cycle ou gestation) (Monniaux et al., 2009). Chez la vache, le cycle comporte le plus souvent 2, voire 3 vagues apparaissant respectivement aux jours 2 et 11 ou aux jours 2, 9 et 16 du cycle respectivement (figure 7). Au cours de chaque vague, on assiste à l'émergence de plusieurs follicules de diamètre supérieur ou égal à 4 mm, parmi lesquels apparaîtra le follicule préovulatoire issu de la dernière vague (Ginther et al., 2013).
FollicleDiameter(mm) 11 Figure 7 : Vagues de croissances folliculaires chez la
vache Au cours de la période pré-pubertaire, la croissance folliculaire se déroule sous la forme des vagues ; cependant, tous les follicules sont anovulatoires à cause d'une insuffisance de l'hormone LH (Savio et al., 1988). Des études ont également montré l'émergence des vagues de croissance folliculaire pendant la gestation mais sans phénomènes de sélection ni de dominance (Ginther et al., 1989). b) Notions et mécanismes de recrutement, sélection et dominance Chaque vague de croissance folliculaire comprend trois phases : recrutement, sélection et dominance (figure 8).
12 Figure 8 : Notion de recrutement, sélection et
dominance d'un follicule chez la vache Lors de la première phase (recrutement), 2 à 3 follicules de diamètre compris entre 3 et 6 mm émergent d'un groupe de follicules tertiaires et deviennent dépendante de l'hormone FSH (Driancourt et al., 1991). La sélection est l'émergence du ou (des) follicule(s) ovulatoire(s) parmi les follicules recrutés. Ceux-ci commencent à sécréter des oestrogènes et de l'inhibine ; ces hormones exercent un rétrocontrôle négatif sur l'hypophyse, abaissant ainsi la sécrétion de la FSH à un niveau inférieur aux besoins folliculaires. A l'exception du ou (des) follicule(s) sélectionné(s) capable(s) de se développer en présence d'un faible taux de FSH, les autres follicules entrent en atrésie (Driancourt et al., 1991). Pendant la dernière phase, le follicule acquiert les récepteurs de LH dans ses cellules de la granulosa et devient dominant (Monniaux et al., 2009). La dominance est à la fois morphologique et fonctionnelle. Elle est qualifiée de morphologique parce qu'elle est exercée par le plus gros follicule présent sur l'un ou l'autre ovaire. Elle est fonctionnelle parce que le follicule dominant est le seul qui soit capable de provoquer la régression des follicules en croissance et d'inhiber le recrutement de nouveaux follicules (Sirois et Fortune, 1988) et d'ovuler dans un environnement hormonal approprié. Le follicule dominant cohabite sur le même ovaire avec 2 à 4 follicules de diamètre compris entre 4 et 8 mm et environ 20 à 80 autres follicules de diamètre compris entre 1 et 3 mm. Cliniquement, la présence sur les 13 ovaires de plus de 10 follicules de diamètre compris entre 3 et 8 mm permet d'exclure celle d'un follicule fonctionnellement dominant. II.2.1.1.3. Régulation de la croissance folliculaire Chez les mammifères, il existe deux phases successives dans la croissance des follicules : la folliculogenèse basale et la folliculogenèse terminale (Monniaux et al., 2009).
Elle est strictement dépendante de la présence de FSH et, pour les stades terminaux de maturation du follicule pré-ovulatoire, de la présence de LH. Elle débute chez la vache lorsque les follicules ont atteint un diamètre de 3 à 4 mm. L'apparition de récepteurs de LH sur les cellules de granulosa est la «signature» d'une maturité complète du follicule, qui devient apte à ovuler. C'est au cours de cette phase que s'effectue la sélection du ou des follicule(s) destiné(s) à ovuler, grâce à un ensemble de mécanismes dont la finalité biologique est de réguler le nombre d'ovulations caractéristique de chaque espèce et de chaque race (Monniaux et al., 2009). II.2.1.2. Atrésie folliculaire Encore appelée involution folliculaire, elle constitue le devenir de la majorité (99.9 %) des follicules présents dans l'ovaire des mammifères. Elle joue donc indirectement un rôle important dans la régulation du taux d'ovulation. Cytologiquement, elle n'est identifiable que chez les follicules antraux par la mise en évidence de pycnose (grains de chromatine 14 condensée) ou d'apoptose (corps apoptotiques) dans les cellules de la granulosa des follicules à antrum ou par l'identification de processus dégénératifs (opacification) au niveau de l'ovocyte. Biochimiquement, elle s'accompagne d'une augmentation des concentrations en enzymes lysosomales et en glycosaminoglycanes ainsi que d'une diminution des concentrations en oestradiol (Monniaux et al., 1999). II.2.1.3. Ovulation et maturation du complexe cumulus-ovocyte II.2.1.3. 1. OvulationL'ovulation est un processus complexe au cours duquel sont induites à la fois la reprise de la méiose de l'ovocyte, l'expansion du cumulus, la rupture du pôle apical du follicule et la restructuration tissulaire associée à la différenciation cellulaire nécessaire à la formation du corps jaune. Seuls les follicules qui expriment de nombreux récepteurs de LH à la surface des cellules de la granulosa sont capables d'ovuler en réponse au pic préovulatoire de LH (Monniaux et al., 2009). L'ovulation se produit 27 heures après la décharge de la LH chez la vache (Sartori et Barros, 2011). |
|