V Discussion
Dans cette partie, nous nous attarderons tout d'abord sur les
aspects techniques (authenticité et conservation de l'ADN) qui
étaient les principaux objectifs de cette étude de
faisabilité. Nous apporterons par la suite quelques résultats en
termes de relation de parenté maternelle entre les individus
étudiés.
1. Authenticité des séquences :
conservation de l'ADN et contaminations - Conservation de l'ADN
sur le site de Jau-Dignac :
Au vue des résultats que nous obtenons, la conservation
de l'ADN des individus analysés apparaît très
satisfaisante. Les résultats génétiques obtenus sont
toutefois fragmentaires (tabl.9). Les neufs individus exhumés
présentaient un état de conservation macroscopique relativement
bon. Cette dernière information conforte bien l'idée qu'il existe
une corrélation entre la conservation macroscopique des restes et leur
conservation moléculaire, comme cela a été décrit
dans la littérature (Cipollaro et al. 1998).
Le caractère récent des
prélèvements ainsi que la mise en congélation
immédiate des échantillons à - 20°C sont deux
facteurs qui ont dû jouer un rôle important de frein à la
dégradation de l'ADN des individus ; ceci pourrait entre autre expliquer
la qualité des résultats obtenus. En effet, il a
été démontré dans la littérature qu'il est
préférable d'analyser un échantillon juste après
son prélèvement sinon les processus de dégradation se
remettent en route (Pruvost et al. 2008). La congélation des
échantillons s'avère également être une
précaution essentielle aux analyses paléogénétiques
puisque celle-ci permet de limiter voir stopper la dégradation de l'ADN
(Barriel 1997). Cette mesure, reconnue depuis longtemps en médecine
légale, n'est cependant pas souvent prise lors d'études portant
sur l'ADN ancien. En effet, comme nous l'avons vu dans la synthèse
bibliographique, les études de (Cappellini et al. 2003 ; Dudar
et al. 2003 ; Rudbeck et al. 2005 ; Adachi et al.
2004) portent sur des vestiges récemment fouillés mais qui n'ont
pas été mis en congélation des leurs
prélèvements.
Les études portant sur des échantillons
prélevés anciennement comme les articles de Bouwman et
al. (2008) et Mooder et al. (2005) n'ont malheureusement pas non
plus bénéficié de cette précaution, ce qui est
encore plus regrettable.
L'état de conservation de l'ADN est souvent
associé à un autre phénomène qui est la
contamination des échantillons par de l'ADN moderne. En effet, plus
l'ADN est dégradé plus
les contaminations exogènes présentes
apparaîtront lors des résultats. Il convient donc, afin de
réaliser des analyses correctes, d'éviter au maximum ces
phénomènes de contamination mais également de pouvoir les
repérer.
- Contamination des échantillons
Comme nous l'avons dit précédemment, les
prélèvements de mandibule n'ont pas tous été
réalisés dans les mêmes conditions.
La sépulture 170 est la seule à avoir
bénéficié de précautions particulièrement
rigoureuses (port de gant, charlotte, mise en sachet puis congélation
immédiate). Les deux autres sépultures, quant à elles,
n'ont pas fait l'objet de port de gants. Toutefois, les autres
précautions ont été prises.
L'analyse de la séquence du fouilleur n°2 (seul
individu ayant prélevé les échantillons des
sépultures 169 et 171) nous permet d'affirmer que cet individu n'a pas
contaminé les échantillons. En effet, son unique mutation en
position 16362 n'a jamais été retrouvée seule dans le
cadre de cette étude. Les quatre autres fouilleurs, qui n'ont jamais
été en contact direct avec les échantillons,
présentent également des séquences absentes des analyses,
ils n'ont donc pas contaminé les échantillons. En
conséquence, l'absence de port de gant lors des
prélèvements des sépultures 169 et 171 n'a eu aucune
conséquence sur les résultats de ces analyses et le fait de
manipuler mains nues, de façon brève, un échantillon ne
s'avère pas un geste contaminant. Cette information confirme que la
manipulation des échantillons à mains nues n'est pas le geste le
plus contaminant, comme précédemment évoqué dans la
littérature (Yang et al. 2005 ; Deguilloux et al.
2009). Cependant, il convient de signaler que les extractions ADN ont
été réalisées à partir d'échantillons
très particuliers (dents intactes en place) et que cette information ne
sera peut-être pas valable pour des prélèvements d'os
(Deguilloux et al. 2009).
Les contaminations observées lors de nos analyses sont
alors issues d'autres sources (réactifs, séquences
préalablement analysées au laboratoire, manipulateurs du
laboratoire).
La séquence associant les mutations 16223 CT, 16256 CT
et 16270 retrouvées dans les derniers témoins a été
identifiée et répertoriée au labo comme contaminante des
réactifs PCR. D'autres contaminations de ce type
(précédemment répertoriées) n'ont pas
été mises en évidence. Aucune contamination via les
manipulateurs du laboratoire n'a été détectée.
Deux séquences comprenant, la mutation 16220 AG pour
l'une et la mutation 16258 AG pour l'autre, ont été
retrouvées dans plusieurs témoins. Selon les études
menées au labo, ces
contaminants proviendraient des lots de primers. Enfin, la
séquence comportant la mutation 16298 TC observée dans les
témoins aérosol est également à répertorier
comme séquence contaminante retrouvée au sein du laboratoire.
Comme nous l'avons montré dans le chapitre
synthèse bibliographique, beaucoup d'études publiées
portant sur l'ADN ancien ne respectent pas toutes les précautions
nécessaires afin de limiter au maximum la dégradation de l'ADN
ainsi que les contaminations par de l'ADN moderne (précautions lors des
prélèvements, congélation des échantillons,
multiples témoins, répétitions dans quelques cas,
clonage). Toutes ces mesures ont été prises dans le cadre de
cette étude de faisabilité ce qui explique sans doute la
qualité des résultats obtenus.
Pour résumer, l'ADN mitochondrial semble globalement
bien conservé et aucune contamination par des fouilleurs et
manipulateurs n'a été mise en évidence. Par
conséquent, des analyses complémentaires peuvent être
envisagées. Dans un premier temps, les résultats obtenus dans le
cadre de cette étude de faisabilité devront être
authentifiés par des répétitions (autres extractions et
amplifications). Si ces nouvelles analyses s'avèrent fructueuses, nous
pourrons nous tourner vers l'ADN nucléaire avec l'analyse des STR. Comme
nous l'avons dit précédemment, ces marqueurs permettront
d'identifier des relations paternelles et de type parent-enfant.
2. Haplogroupes retrouvés
Ces analyses préliminaires ont permis de
caractériser deux haplogroupes européens différents J et X
chez trois individus au moins. La mise en évidence de mutations
diagnostiques dans ces séquences anciennes nous laisse confiants quant
à l'attribution de ces haplogroupes.
Pour deux individus (présentant la même
séquence), il n'a pas été possible de préciser
l'haplogroupe à partir des séquences HVRI seules, mais il
pourrait s'agir de l'haplogroupe H ou K, tous deux européens.
- L'haplogroupe J
La sépulture 169 présente deux individus
d'haplogroupe J mais d'haplotypes différents.
Celui-ci pourrait également être présent
chez le 3ème inhumé de la sépulture 171 mais
des analyses complémentaires des fragments A et B seraient
nécessaires afin de le confirmer.
Cet haplogroupe J serait apparu au Moyen Orient il y a environ
45 000 ans. Il serait aujourd'hui majoritairement européen. Le graphique
ci-dessous (fig.10) présente les fréquences de cet haplogroupe
retrouvées en Europe de nos jours (Logan 2009).
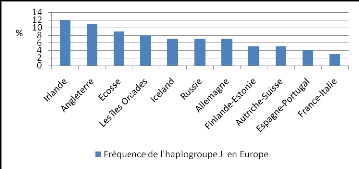
Figure 10 : Fréquence actuelle de
l'haplogroupe J en Europe d'après Logan (2009).
Une étude phylogéographique menée en
France montre que l'haplogroupe J serait aujourd'hui essentiellement
retrouvé à l'Ouest. La fréquence la plus importante de
celui-ci serait observée en Bretagne (Finistère) avec 9%. On le
retrouverait également en Centre Ouest (Corrèze, Dordogne), ou il
serait un peu moins présent, avec une fréquence de 4% (Dubut
2004). Il convient toutefois d'ajouter que les estimations de cette
étude sont très limitées du fait du très faible
échantillonnage réalisé, concernant seulement 3
régions françaises et 50 individus, ces informations sont donc
à prendre avec précautions.
Le blast de séquences endogènes obtenues pour le
troisième individu nous renvoie à plusieurs études dans
lesquelles sont retrouvées des séquences homologues à
100%. Ces séquences sont retrouvées en Europe du Nord
(Royaume-Uni, Islande) ainsi qu'en Hongrie.
- Haplogroupe H ou K ?
La mutation 16224 TC retrouvée chez les deux premiers
inhumés de la sépulture 170 renvoie aux deux haplogroupes K et H
qui sont tous deux majoritairement observés en Europe de l'Ouest et du
Nord (Richards 2000). En Europe, l'haplogroupe K, apparu il y a environ 50 000
ans, est majoritairement présent en Scandinavie. En France, celui-ci est
plus particulièrement observé en Bretagne ainsi qu'en
Centre-Ouest (Dubut 2004). Cependant, là
aussi ces estimations sont limitées par le faible
échantillon concerné. L'haplogroupe H, quant à lui, est
l'haplogroupe le plus présent en Europe actuellement avec une
fréquence variant de 35 à 55 % suivant les pays et les
régions (Richards 2000).
- L'haplogroupe X
L'haplogroupe X retrouvé chez le 1er
inhumé de la sépulture 171 est un haplogroupe assez rare mais
présent dans beaucoup de région du monde. L'haplogroupe X dans
son ensemble représente environ 2 % de la population d'Europe, du
Proche-Orient et d'Afrique du Nord. Le sous-groupe X2,
spécifiquement européen et à priori absent de la majeure
partie de l'Asie continental mais se retrouve également parmi les cinq
haplogroupes majeurs des populations indigènes d'Amérique. Le
graphique ci-dessous présente la fréquence de cet haplogroupe en
Europe (fig.11) (Logan 2009).
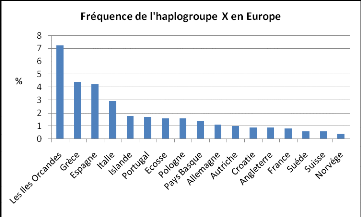
Figure 11 : Fréquence de l'haplogroupe X en
Europe d'après Logan (2009).
En France, l'haplogroupe X est uniquement présent dans
le Nord (Bretagne et Nord-Pas de calais) à une fréquence
d'environ 0,4% (Dubut 2004). Ces estimations se basent sur un
échantillon très réduit, elles n'ont donc qu'une valeur
indicative et très approximative.
Les mutations présentes chez le 1er
inhumé du sarcophage 171 ne permettent pas d'obtenir un haplotype
précis car celles-ci sont présentes au sein des deux sous groupes
X1 et X2. D'autres régions de l'ADN mt seraient nécessaires afin
de pouvoir acquérir cette information.
- Cas des deux derniers inhumés de la sépulture
171
La détermination d'haplogroupe chez les deux derniers
inhumés de la sépulture 171 n'a pas été possible.
Comme nous l'avons dit précédemment, l'hypothèse selon
laquelle le dernier inhumé de ce sarcophage serait d'haplogroupe J1B1
par l'association des mutations 16069 CT, 16126 TC, 16145 CT, 16222 CT et 16261
CT serait également envisageable. On resterait alors dans une
continuité d'haplogroupe J. Des analyses complémentaires portant
sur les fragments A et B seraient nécessaires afin de clarifier cela.
En résumé, nous obtenons dans le cadre de cette
étude, des haplogroupes s'intégrant parfaitement dans le pool
génétique européen, peut-être plus
particulièrement du Nord-ouest du continent. Ceci paraît donc tout
à fait logique et attendu et cela pourrait être un argument
supplémentaire d'authenticité des séquences obtenues.
3. Liens de parentés
Comme nous l'avons dit précédemment, les
séquences obtenues pour les deux premiers inhumés de la
sépulture 170 paraissent authentiques. On observe l'unique mutation
16224 TC chez ces deux individus sur deux ou trois fragments. On peut donc
d'ores et déjà penser que ces deux individus sont
apparentés du coté maternel.
Cependant, la similitude génétique de ces deux
individus pourrait être expliquée par une deuxième
éventualité, jusque là non abordée dans la
littérature. En effet, les os de la 1ere inhumée auraient pu
être contaminés par les « jus organiques » produits par
la deuxième inhumée lors de sa décomposition. Cette
hypothèse ne peut pas être exclue irrévocablement,
néanmoins, la qualité des prélèvements (dents
intactes et en place) ne permet pas d'envisager sérieusement cette
possibilité. La logique privilégie donc l'hypothèse selon
laquelle les deux individus seraient bien apparentés maternellement.
Les deux individus, une adolescente de sexe féminin
ainsi qu'une femme âgée entre 20 et 30 ans, pourraient avoir eu
des relations de plusieurs types, tel que mère-fille, soeurs, tante-
nièce ou encore des relations bien plus éloignées
(fig.12).
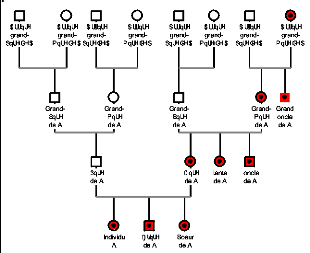
Figure 12 : Arbre généalogique
montrant quels individus sont issus d'une même lignée
mitochondriale. Les individus en rouge présentent le même
haplogroupe mitochondriale que l'individu A.
Les méthodes archéothanalogiques
pratiquées sur le chantier ont démontré que l'adolescente
aurait été inhumée en 1er. Puis ces os auraient
été réduits afin de fournir de l'espace à l'autre
jeune femme. Malheureusement, il n'existe aucun argument taphonomique
permettant d'obtenir un ordre de grandeur quant au temps séparant les
deux inhumations. La décomposition du corps peut s'être faite
aussi bien en quelques mois qu'en quelques années. Il n'est donc pas
possible d'affiner nos hypothèses concernant le type de lien unissant
ces deux femmes. Des analyses STR pourraient permettre de mettre en
évidence des relations de type mère-fille ou soeurs, si cela est
le cas.
La présence du périnatal dans ce sarcophage
reste toujours un mystère, puisque malheureusement les analyses
génétiques n'ont rien fournit pour cet individu.
À cette époque, les enfants n'étaient
baptisés qu'à l'âge de 3 ou 4 ans, lorsqu'on était
certain que l'enfant était en bonne santé et allait vivre, car un
baptême coûtait fort cher. Les enfants décédés
sans être baptisés étaient donc enterrés en tant que
non-chrétiens, hors de l'enclos sacré. Il se pose alors toujours
la question de la présence de ce périnatal au sein de cette
nécropole et surtout au sein d'un sarcophage. L'hypothèse la plus
plausible expliquant cette pratique est que ce bébé aurait
été déposé aux coté de l'un de ses parents.
Au vu des arguments archéothanatologiques énoncés dans la
première partie du mémoire, ce périnatal aurait pu
être l'enfant de la femme (2nde inhumée) située
à ses cotés. Si tel était le cas ce sarcophage
regrouperait donc trois personnes issues d'une même famille.
Cette étude a permis le rapprochement biologique de
deux individus seulement mais certains résultats fragmentaires de ces
analyses ne permettent pas d'écarter catégoriquement d'autres
liens de parenté maternels entre d'autres individus, au sein des
sarcophages et entre les sarcophages. Toutefois, au sein des tombes, nous
pouvons d'ores et déjà exclure tous liens de parenté
maternels entre les premier et troisième inhumés de la
sépulture 169 ainsi qu'entre les premier et troisième
inhumé de la sépulture 171 (fig.13). Nous pouvons donc aussi
écarter tous lien de parenté entre ces quatre individus.
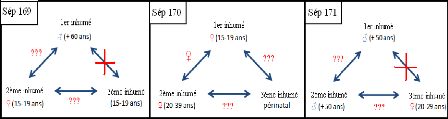
Figure 13 : Informations acquises en termes de
relations de parenté maternelles.
X : aucun lien de parenté maternel ; ??? : Pas de
liens maternels démontrés ; ? : liens maternels
démontrés.
Ces analyses paléogénétiques très
préliminaires permettent de mettre en évidence 2 cas de figures
distincts :
Nous retrouvons des individus apparentés maternellement
déposés dans le même sarcophage mais aussi des individus
non apparentés maternellement déposés dans le même
sarcophage. Cette information nous montre que l'on ne peut pas forcément
globaliser les résultats. En effet, la mise en évidence de
relations de parenté au sein d'un sarcophage ne veut pas dire que cela
est forcément le cas pour le sarcophage voisin. Nous sommes donc peut
être face à des gestes funéraires différents.
Ces dernières informations posent la question de
l'apport des données paléogénétiques : sont-elles
potentiellement généralisables à partir d'un site, d'un
sarcophage, d'un groupe dans la nécropole ? La synthèse
bibliographique présentée précédemment nous montre
bien que dans la plupart des sites étudiés, il n'est pas possible
de généraliser les phénomènes de regroupements
familiaux car ceux-ci sont souvent propres à une seule sépulture
ou à un secteur particulier.
| 

