1.2. Analyses quantitatives
1.2.1. Analyse par années
1.2.1.1. Fréquences annuelles de toutes les
espèces
Les fréquences de toutes les espèces biomonitrices
sont données par la figure 9.
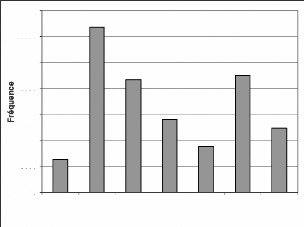
25000
20000
35000
30000
15000
10000
5000
0
1999 2000 2001 2002 2003 2004 2005
Année
Figure 9 : Evolution des fréquences de toutes les
espèces biomonitrices
Les fréquences les plus élevées sont
celles de 2000 et de 2001. L'année 2003 a la plus faible
fréquence. L'évolution de ces fréquences montre une forte
augmentation de 1999 à 2000 puis une baisse de 2001 jusque en 2003. Il y
a une nouvelle hausse en 2004 puis une chute en 2005.
La faible fréquence de 1999 s'expliquerait par la mise
en place du système ; et donc sa non maîtrise. La fréquence
très élevée de 2000 s'expliquerait par deux raisons
essentielles. D'abord l'engouement que les collecteurs de données
avaient à bien travailler, ensuite la surestimation des effectifs des
groupes de singes (PNT, 2002) qui représentent 58% à 72% des
effectifs (Tableau A1). La moyenne des groupes de singes était de 16,25
individus en cette année.
La baisse de 2001 à 2002 serait le fait de la
réadaptation de la méthode avec une estimation des effectifs
revue à la baisse (PNT, 2002). Les groupes de singes faisant en moyenne
11,07 individus en 2001. Cette baisse se justifierait aussi par le manque
d'engouement des collecteurs de données au travail. Ces derniers ne
percevaient pas régulièrement leur salaire du fait de la fin de
la première phase du projet PACPNT. Pour ce qui est de 2003, la faible
fréquence est due en grande partie à la guerre qu'a connue la
Côte-d'Ivoire en septembre 2002. La guerre a causé l'interruption
des travaux dans tous les secteurs durant les six
premiers mois de l'année 2003. A cela, s'ajoute le taux
de braconnage qui aurait augmenté après l'arrêt des
hostilités et avant la reprise des travaux de surveillance (N'Goran,
2004). Cela a sûrement rendu les animaux tels que les singes et les
antilopes plus craintifs.
L'arrêt des hostilités en Côte-d'Ivoire et
la reprise des activités de surveillance et de collecte des
données ont permis de rehausser le niveau en 2004. La fréquence
peu élevée de 2005 s'explique par le fait que les travaux n'ont
porté que sur la moitié de l'année (janvier à
juin).
1.2.1.2. Comparaison des années pour toutes les
espèces
Le test de Levene (Tableau A3) ayant montré que la
distribution de la population ne suit pas une loi normale, nous avons fait un
test de Kruskal-Wallis (Tableau A4). La comparaison des années a
montré une différence significative entre elles (p = 0,000). Cela
nous a conduit à faire un test de ségrégation des moyennes
pour situer les différences significatives (Tableau 4).
Tableau 4 : Comparaisons des taux d'observations des
années par paire
|
(I) ANNÉE
|
(J) ANNÉE
|
Différence des
moyennes (I-J)
|
Erreur
standard
|
Signification
|
|
1999
|
2000
|
-1,651(*)
|
0,1507
|
0,000
|
|
2001
|
-,838(*)
|
0,1158
|
0,000
|
|
2002
|
-,349(*)
|
0,1142
|
0,047
|
|
2003
|
-0,284
|
0,1097
|
0,184
|
|
2004
|
-,366(*)
|
0,0967
|
0,003
|
|
2005
|
-,522(*)
|
0,1097
|
0,000
|
|
2000
|
2001
|
,813(*)
|
0,1471
|
0,000
|
|
2002
|
1,302(*)
|
0,1459
|
0,000
|
|
2003
|
1,367(*)
|
0,1423
|
0,000
|
|
2004
|
1,285(*)
|
0,1326
|
0,000
|
|
2005
|
1,129(*)
|
0,1424
|
0,000
|
|
2001
|
2002
|
,489(*)
|
0,1095
|
0,000
|
|
2003
|
,554(*)
|
0,1047
|
0,000
|
|
2004
|
,472(*)
|
0,09 10
|
0,000
|
|
2005
|
0,316
|
0,1047
|
0,052
|
|
2002
|
2003
|
0,065
|
0,1030
|
1,000
|
|
2004
|
-0,017
|
0,0890
|
1,000
|
|
2005
|
-0,173
|
0,1030
|
0,871
|
|
2003
|
2004
|
-0,081
|
0,0831
|
1,000
|
|
2005
|
-0,238
|
0,0979
|
0,274
|
|
2004
|
2005
|
-0,156
|
0,0831
|
0,726
|
* différence significative au seuil 5%
Les comparaisons appariées des moyennes des taux
d'observations montrent bien que les
années 1999, 2000 et 2001 sont
significativement différentes des autres années mais
également entre elles. En revanche, les différences
entre 2001 et 2005, entre 1999 et 2003 ne sont pas significatives. Il en est de
même pour les années 2002, 2003, 2004 et 2005.
Ceci dit, nous allons porter un regard sur les espèces les
plus braconnées que sont les singes et les antilopes.
1.2.1.3. Fréquences annuelles des singes et
antilopes
A ce niveau, nous avons considéré les
espèces les plus abondantes. Les figures 10 et 11 donnent les
espèces dominantes de chacun de ces groupes d'espèces. Les singes
sont dominés par Cercopithecus diana diana (34,93%) et
Piliocolobus badius badius (23,04%). Quant aux antilopes, elles sont
fortement dominées par Cephalophus maxwelli (82,84).
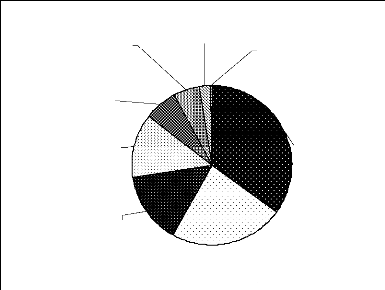
Cercopithecus
mona; 6,59%
Cercopithecus
petaurista;
13,07%
Colobus
polykomos
polykomos;
5,39%
Cercocebus
atys atys;
14,48%
Colobus verus;
2,29%
Piliocolobus
badius badius;
23,04%
Cercopithecus
nictitans;
0,20%
Cercopithecus
diana diana;
34,93%
Figure 10 : Répartition des fréquences
des singes entre les espèces

Neotra gus
pygmaeus;
2,65%
Cephalophus
olgilby;
3,14%
Cephalophus
niger; 5,60%
Cephalophus zebra; 2,61%
Hyemoschus
aquaticus;
1,82%
Tragelaphus
scriptus
scriptus;
1,02%
Cephalophus
silviculto r;
0,61%
Cephalophus
jentenki;
0,72%
Cephalophus
maxwelli;
8 1,84%
Figure 11 : Répartition des fréquences
des antilopes entre les espèces
Les fréquences de ces espèces sont
présentées par les figures 12, 13 et 14.
Les allures sont similaires à l'évolution
donnée par toutes les espèces regroupées (Figure 9). Cette
similarité s'explique par les raisons évoquées dans le cas
de toutes les espèces. A cela s'ajoute le fait que C. diana diana
et P. badius badius sont les espèces les plus
dominantes.
|
9000
8000
7000
6000
5000
4000
3000
2000
1000
0
|
|
1999 2000 2001 2002 2003 2004 2005
Année
Figure 12 : Evolution des fréquences de
Cercopithecus diana diana
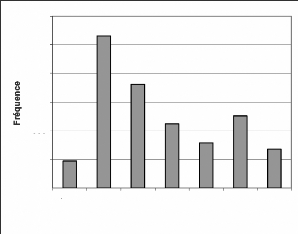
4000
2000
6000
5000
3000
1000
0
1999 2000 2001 2002 2003 2004 2005
Année
Figure 13 : Evolution des fréquences de
Piliocolobus badius badius
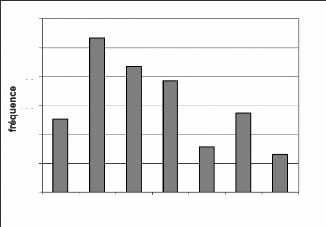
400
600
500
300
200
100
0
1999 2000 2001 2002 2003 2004 2005
Année
Figure 14 : Evolution des fréquences de
Cephalophus maxwelli
1.2.1.4. Comparaison des années pour les singes et
antilopes
La comparaison des années s'est faite sur la base des
taux d'observation. Ces taux ont porté sur les trois espèces
sus-considérées. Ici également, le test
d'homoscédasticité (Tableau A3) a montré que les
populations ne sont pas normalement distribuées (p=4,551
E-19) . Nous avons donc fait un test de Kruskal-Wallis (Tableau A4).
Ce test a montré des différences significatives (p=0,04) entre
les années pour chacune des trois espèces. Pour mettre en
exergue les années qui sont significativement
différentes, nous avons fait des comparaisons appariées (Tableaux
5, 6 et 7).
Tableau 5 : Comparaison par paires des taux
d'observations de Cercopithecus diana
diana des
années
|
(I) ANNÉE
|
(J) ANNÉE
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
2000
|
-4,8479(*)
|
0,8499 1
|
0,000
|
|
2001
|
-1,576
|
0,6405
|
0,267
|
|
2002
|
0,4825
|
0,5542
|
1,000
|
|
1999
|
|
|
|
|
|
2003
|
0,1275
|
0,58999
|
1,000
|
|
2004
|
-0,1468
|
0,53936
|
1,000
|
|
2005
|
-0,8211
|
0,61206
|
0,985
|
|
2001
|
3,2719(*)
|
0,78768
|
0,001
|
|
2002
|
5,3304(*)
|
0,71927
|
0,000
|
|
2000
|
2003
|
4,9754(*)
|
0,74719
|
0,000
|
|
2004
|
4,7011(*)
|
0,70789
|
0,000
|
|
2005
|
4,0269(*)
|
0,76473
|
0,000
|
|
2002
|
2,0585(*)
|
0,45303
|
0,000
|
|
2001
|
2003
|
1,7035(*)
|
0,49617
|
0,014
|
|
2004
|
1 ,4292(*)
|
0,43474
|
0,023
|
|
2005
|
0,7549
|
0,52222
|
0,966
|
|
2003
|
-0,355
|
0,37827
|
1,000
|
|
2002
|
2004
|
-0,6293
|
0,29309
|
0,498
|
|
2005
|
-1,3036(*)
|
0,41183
|
0,036
|
|
2003
|
2004
|
-0,2742
|
0,35616
|
1,000
|
|
2005
|
-0,9485
|
0,45887
|
0,574
|
|
2004
|
2005
|
-0,6743
|
0,39163
|
0,851
|
* différence significative au seuil 5%
Tableau 6 : Comparaison par paires des taux
d'observations de Piliocolobus badius
badius des
années
|
(I) ANNÉE
|
(J) ANNÉE
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
2000
|
-4,4568(*)
|
1,8240
|
0,0287
|
|
2001
|
0,5869
|
1,3104
|
1,000
|
|
1999
|
2002
|
2,9956
|
1,2062
|
0,310
|
|
2003
|
2,3302
|
1,2472
|
0,775
|
|
2004
|
2,4502
|
1,1998
|
0,646
|
|
2005
|
2,4479
|
1,2502
|
0,706
|
|
2001
|
5,0437(*)
|
1,5416
|
0,027
|
|
2002
|
7,4524(*)
|
1,4541
|
0,000
|
|
2000
|
2003
|
6,7871(*)
|
1,4882
|
0,000
|
|
2004
|
6,9071(*)
|
1,4488
|
0,000
|
|
2005
|
6,9048(*)
|
1,4908
|
0,000
|
|
2002
|
2,4087(*)
|
0,7103
|
0,017
|
|
2001
|
2003
|
1,7434
|
0,7778
|
0,425
|
|
2004
|
1,8634
|
0,6994
|
0,160
|
|
2005
|
1,861
|
0,7827
|
0,322
|
|
2003
|
-0,6653
|
0,5855
|
0,998
|
|
2002
|
2004
|
-0,5453
|
0,4765
|
0,998
|
|
2005
|
-0,5477
|
0,5921
|
1,000
|
|
2003
|
2004
|
0,1200
|
0,5722
|
1,000
|
|
2005
|
0,1177
|
0,6715
|
1,000
|
|
2004
|
2005
|
-0,0023
|
0,5789
|
1,000
|
* différence significative au seuil 5%
Tableau 7 : Comparaison par paires des taux
d'observations de Cephalophus maxweii
des années
|
(I) ANNÉE
|
(J) ANNÉE
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
2000
|
-,1851(*)
|
0,0496
|
0,005
|
|
2001
|
-0,1442
|
0,0482
|
0,061
|
|
1999
|
2002
|
-0,0928
|
0,0573
|
0,906
|
|
2003
|
0,0372
|
0,0425
|
1,000
|
|
2004
|
0,012
|
0,0420
|
1,000
|
|
2005
|
0,0182
|
0,0440
|
1,000
|
|
2001
|
0,0409
|
0,0558
|
1,000
|
|
2002
|
0,0923
|
0,0638
|
0,966
|
|
2000
|
2003
|
,2222(*)
|
0,0510
|
0,000
|
|
2004
|
,1971(*)
|
0,0506
|
0,002
|
|
2005
|
,2033(*)
|
0,0522
|
0,003
|
|
2002
|
0,0514
|
0,0627
|
1,000
|
|
2001
|
2003
|
,1813(*)
|
0,0496
|
0,007
|
|
2004
|
,1562(*)
|
0,0492
|
0,034
|
|
2005
|
,1624(*)
|
0,0509
|
0,034
|
|
2003
|
0,1299
|
0,0585
|
0,439
|
|
2002
|
2004
|
0,1048
|
0,0581
|
0,794
|
|
2005
|
0,111
|
0,0596
|
0,748
|
|
2003
|
2004
|
-0,0251
|
0,0436
|
1,000
|
|
2005
|
-0,0189
|
0,0455
|
1,000
|
|
2004
|
2005
|
0,0062
|
0,0451
|
1,000
|
* différence significative au seuil 5%
Ces comparaisons donnent, au niveau des deux espèces de
singes, des résultats presque identiques à ceux de toutes les
espèces regroupées. Cependant, il y a des variations importantes
avec C. maxwelli. Pour cette espèce, les différences ne
sont pas significatives entre l'année 2002 et toutes les autres
années. Il en est de même des différences entre les
années 1999, 2003, 2004 et 2005. En revanche, les années 2000 et
2001 diffèrent significativement du reste des autres années.
Cette situation serait provoquée par la guerre qui avait favorisé
le braconnage en 2003 et dont les conséquences se font ressentir sur les
autres années.
1.2.2. Analyse par secteur
1.2.2.1. Fréquences des secteurs pour toutes les
espèces
Les fréquences des secteurs sont données par la
Figure 15.
Les animaux sont plus nombreux dans les secteurs IV et III. Les
secteurs I et II ont des
fréquences très faibles. L'abondance
des animaux dans les secteurs III et IV s'expliquerait par
la
présence très marquée des singes ; mais également
par la stabilité de ces zones (braconnage
peu intense). Les très faibles fréquences des
secteurs I et II s'expliqueraient par les effets du braconnage intense. Les
animaux de ces zones sont craintifs et peu nombreux (Newing, 1994 ;
Hoppe-Dominik, 1998 ; Gerhard, 2004). Ils ont sûrement migrés vers
des zones plus stables. Le braconnage peut être à la base de la
faible fréquence du secteur V (Hoppe-Dominik, 1995)
|
25000
20000
15000
10000
5000
0
|
|
I II III IV V
Secteur
Figure 15: Répartition des fréquences de
toutes les espèces biomonitrices par secteur
1.2.2.2. Comparaison des secteurs pour toutes les
espèces
La comparaison des secteurs par le test de Kruskal-Wallis
montre qu'il y a une différence significative entre eux (Tableau A4).
Une comparaison appariée de ces secteurs (Tableau 8) montre qu'ils sont
significativement différents les uns des autres.
Tableau 8 : Comparaisons par paires des taux
d'observations des secteurs
|
(I)
SECTEUR
|
(J)
SECTEUR
|
Différence des
moyennes (I-J)
|
Erreur
standard
|
Signification
|
|
I
|
II
|
0,191(*)
|
0,0602
|
0,015
|
|
III
|
-0,798(*)
|
0,0585
|
0,000
|
|
IV
|
-2,822(*)
|
0,1039
|
0,000
|
|
V
|
-0,132(*)
|
0,0468
|
0,047
|
|
II
|
III
|
-0,988(*)
|
0,06 13
|
0,000
|
|
IV
|
-3,013(*)
|
0,1056
|
0,000
|
|
V
|
-0,323(*)
|
0,0504
|
0,000
|
|
III
|
IV
|
-2,025(*)
|
0,1046
|
0,000
|
|
V
|
0,665(*)
|
0,0483
|
0,000
|
|
IV
|
V
|
2,690(*)
|
0,0986
|
0,000
|
* différence significative au seuil de 5%
Comme dans le cas des années, nous avons ouvert une
lucarne sur les singes et les antilopes pour avoir les tendances
spécifiques.
1.2.2.3. Fréquences des singes et des antilopes
dans les secteurs
La répartition des fréquences des trois
espèces sont données par les Figures 16, 17 et 18.
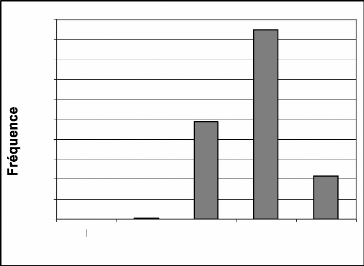
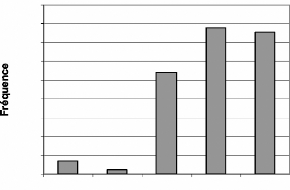
Les singes ne sont presque pas rencontrés dans les
secteurs I et II. Ils sont en revanche très nombreux dans le secteur IV.
C. badius se rencontre presque exclusivement dans le secteur IV. Les
tendances sont identiques avec C. maxwelli. Cependant les
écarts sont moins marqués entre les secteurs III, IV et V.
La répartition de ces fréquences confirme
l'influence du braconnage dans les secteurs I et II. Newing (1994) a
montré que les C. maxwelli avaient une préférence
prononcée pour la forêt primaire (secteurs IV et V) que pour la
forêt secondaire (secteur III).
I II III IV V
Secteur
500
0
5000
4500
4000
3500
3000
2500
2000
1500
1000
Figure 16 : Répartition des fréquences de
Cercopithecus diana diana par secteur
|
6000
5000
4000
3000
2000
1000
0
|
|
I II III IV V
Secteur
Figure 17 : Répartition des fréquences de
Piiocolobus badius badius par secteur
I II III IV V
900
800
700
600
500
400
300
200
100
0
Secteur
Figure 18 : Répartition des fréquences de
Cephalophus maxweii par secteur
1.2.2.4. Comparaison des secteurs pour les singes et
antilopes
La comparaison (Tableau A4) des secteurs a montré qu'il y
a des différences significatives entre eux pour chacune des trois
espèces (p = 3,980e-14).
Les comparaisons appariées (Tableaux 9, 10 et 11)
donnent des résultats presque similaires à celles de toutes les
espèces regroupées. Cependant, les différences entre les
secteurs I et II ne sont pas significatives pour les trois espèces.
Tableau 9: Comparaison par paires des taux d'observations
de Cercopithecus diana diana des secteurs
|
(I)
SECTEUR
|
(J)
SECTEUR
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
II
|
0,7639
|
0,2789
|
0,131
|
|
III
|
-3,2389(*)
|
0,3028
|
0,000
|
|
I
|
IV
|
-7,1048(*)
|
0,4086
|
0,000
|
|
V
|
-1,7042(*)
|
0,2308
|
0,000
|
|
III
|
-4,0028(*)
|
0,2900
|
0,000
|
|
II
|
IV
|
-7,8687(*)
|
0,3992
|
0,000
|
|
V
|
-2,4681(*)
|
0,2136
|
0,000
|
|
IV
|
-3,8659(*)
|
0,4163
|
0,000
|
|
III
|
V
|
1,5347(*)
|
0,2440
|
0,000
|
|
IV
|
V
|
5,4006(*)
|
0,3672
|
0,000
|
* différence significative au seuil 5%
Tableau 10 : Comparaison par paires des taux
d'observations de Piliocolobus badius badius des secteurs
|
(I)
SECTEUR
|
(J)
SECTEUR
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
II
|
-0,3886
|
0,1818
|
0,324
|
|
I
|
III
|
-2,3264(*)
|
0,4278
|
0,000
|
|
IV
|
-8,6064(*)
|
0,4493
|
0,000
|
|
V
|
-1,8949(*)
|
0,1669
|
0,000
|
|
III
|
-1,9378(*)
|
0,4324
|
0,000
|
|
II
|
IV
|
-8,2178(*)
|
0,4537
|
0,000
|
|
V
|
-1,5063(*)
|
0,1783
|
0,000
|
|
III
|
IV
|
-6,2800(*)
|
0,5965
|
0,000
|
|
V
|
0,4315
|
0,4263
|
0,977
|
|
IV V
|
6,7115(*)
|
0,4479
|
0,000
|
* différence significative au seuil 5%
Tableau 11 : Comparaison par paires des taux
d'observations de Cephalophus maxweii
des secteurs
|
(I)
SECTEUR
|
(J)
SECTEUR
|
Différence
des
moyennes
(I-J)
|
Erreur
standard
|
Signification
|
|
II
|
-0,0433
|
0,0484
|
0,992
|
|
I
|
III
|
-,0968(*)
|
0,0315
|
0,024
|
|
IV
|
-,2316(*)
|
0,0309
|
0,000
|
|
V
|
-,4218(*)
|
0,0448
|
0,000
|
|
III
|
-0,0535
|
0,0492
|
0,967
|
|
II
|
IV
|
-,1884(*)
|
0,0489
|
0,009
|
|
V
|
-,3786(*)
|
0,0586
|
0,000
|
|
III
|
IV
|
-,1348(*)
|
0,0321
|
0,000
|
|
V
|
-,3250(*)
|
0,0456
|
0,000
|
|
IV
|
V
|
-,1902(*)
|
0,0452
|
0,000
|
* différence significative au seuil 5%
Les secteurs I et II, situés du côté Est
du PNT, sont soumis aux mêmes pressions de braconnage. Pour ce qui est du
secteur IV, la présence de chercheurs dans la zone a joué en
faveur de celui-ci. Certaines espèces de singes y
bénéficient d'un programme d'habituation à l'homme. Cette
présence y a limité l'intensité du braconnage
(Hoppe-Dominik, 1996).
1.2.3. Densités
Rappelons que pour l'analyse des densités, nous n'avons
considéré que les secteurs III, IV et V. Nous avons tracés
les courbes évolutives des densités de ces secteurs (Figure 19,
20 et 21) pour trois espèces. Il s'agit notamment de C. diana
diana, P. badius badius et C. maxwelli.
1.2.3.1. Densités de Cercopithecus diana
diana
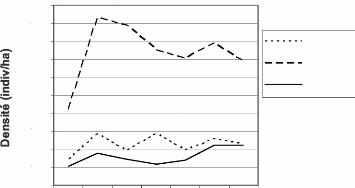
Du point de vue des secteurs, les densités sont plus
élevées dans le secteur IV que dans les deux autres secteurs
(Figure 19). Elles font plus du triple de celle des autres secteurs.
Dans le secteur IV, les densités de C. diana
diana varient entre 2,10 et 4,67 (Tableau A11). Elles sont comprises entre
0,72 et 1,43 dans le secteur V et entre 0,52 et 1,80 dans le secteur III.
Nous obtenons les mêmes tendances que Gerhard (2004).
Cependant il y a quelques des écarts entre les chiffres. En revanche,
nos chiffres sont proches des résultats obtenus par Refisch et
Koné en 2001 dans des zones des secteurs IV et V.
En considérant les années, nous retenons que les
densités évoluent en dents de scie au sein de
tous les
secteurs. Les densités des trois secteurs ont connu une hausse de 1999
à 2000. Par la
suite, elles ont fortement régressées jusqu'en
2003 pour connaître une nouvelle hausse à partir de 2004.
Les difficultés de 2002 auraient eu un impact sur ces
densités. Soit, le manque de surveillance a favorisé un intense
braconnage, soit le manque d'engouement au travail des collecteurs de
données a donné des résultats erronés. Le
braconnage, favorisé par la guerre, expliquerait les faibles
densités de 2003.
1999 2000 2001 2002 2003 2004 2005
Année
Secteur V Secteur IV Secteur III
5
4,5
4
3,5
3
2,5
2
1,5
1
0,5
0
Figure 19 : Evolution des densités de
Cercopithecus diana diana dans
les secteurs III, IV et
V
1.2.3.2. Densités de Piliocolobus badius
badius
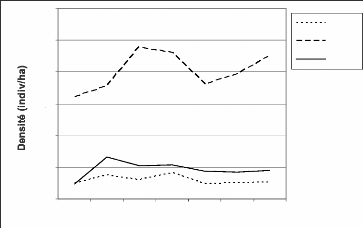
L'évolution des densités de Piliocolobus
badius badius (Figure 20) donne presque les mêmes tendances que
C. diana diana. Le secteur IV est hautement plus dense que les deux
autres secteurs. Les densités des deux autres secteurs sont quasi
stables avec une légère tendance à la baisse depuis 2004.
L'évolution des densités des secteurs traduit l'effet de la
guerre de 2002 sur le PNT.
1999 2000 2001 2002 2003 2004
2005Année
Secteur V
Secteur IV
Secteur III
3
2,5
2
1,5
1
0,5
0
Figure 20 : Evolution des densités de
Piliocolobus badius badius
dans les secteurs III, IV et
V
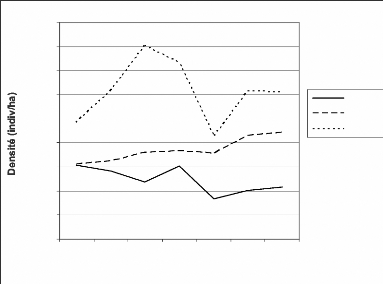
1.2.3.3. Densités de Cephalophus
maxwelli
Les densités de Cephalophus maxwelli (Figure
21) sont plus faibles (0,27) comparativement aux singes. Du point de vu des
secteurs, le secteur V est le plus dense. Comme dans le cas des singes, le
secteur III demeure le moins dense. Les chiffres sont proches de ceux obtenus
par Gerhard en 2004. Les observations indirectes faites par Hoppe-Dominik
(1997) ont donné des résultats proches. Newing (1994) avait
trouvé des densités de 102 individus/km2 dans la
région du secteur IV.
Du point de vu des années, les densités
connaissent des variations. Dans le secteur V, La densité augmente de
1999 à 2001 puis chute. A l'inverse, elle connaît une augmentation
continuelle depuis 1999 dans le secteur IV. Le secteur III, pour sa part, a
connu une baisse graduelle de 1999 à 2002 puis une hausse de 2003
à 2005.
La baisse des densités de 2003 dans les secteurs III et
V serait à l'actif du braconnage. Quant à la hausse dans le
secteur IV, elle peut se traduire par une hausse de la population dans ce
secteur (Hoppe-Dominik, 1999). La rupture de cette croissance en 2003 peut
être l'effet du braconnage favorisé par la guerre de septembre
2002.
0,45
0,35
0,25
0,15
0,05
0,4
0,3
0,2
0,1
0
1999 2000 2001 2002 2003 2004 2005
Année
Secteur III Secteur IV Secteur V
Figure 21 : Evolution des densités de
Cephalophus maxweii dans les secteurs III, IV et V
|
|


