b. les rétrotransposons sans LTR :
Ces éléments sont caractérisés par
une queue poly A ou une région riche en A à leur
extrémité 3' (Finnegan 1992). Ils possèdent à
chaque extrémité de courtes répétitions directes.
La grande diversité des rétrotransposons sans LTR et leur
similitude avec les rétrotransposons à LTR suggèrent que
les rétrotransposons sans LTR seraient à l'origine des
rétrotransposons à LTR (Craig, 2002). La sous-classe des
rétrotransposons sans LTR se divisent en deux familles : les LINE et les
SINE.
b.1 LINE (Long Interspersed Nuclear
Elements):
Les LINE peuvent mesurer 1 à 7 kb et possèdent
généralement deux ORF, correspondant aux gènes gag
et pol. Contrairement aux rétrotransposons à LTR,
le gène pol est dépourvu d'intégrase. La plupart
de ces éléments sont tronqués en 5' et possèdent
des promoteurs internes qui leur permettent d'être transcrits par une ARN
polymérase II (McLean et al. 1993).
Bien qu'ils soient rétrotranscrits, les
éléments LINE ont un système d'intégration
complètement différent des rétrotransposons à LTR.
En effet, la transcriptase inverse reconnaît le transcrit et initie la
rétrotranscription, en même temps que l'intégration au
niveau d'une coupure de l'ADN génomique effectuée par une
endonucléase (Luan et al. 1993).
Il y a plusieurs familles de LINE telles que LINE 1
(L1), L2 et L3. Mais dans le génome humain on
trouve généralement des L1 (figure 2).
· Structure, expression et
répartition des L1.
Un fragment L1 de 6 kb possède une
région non traduite 5' (UTR) de 906 nucléotides, deux phases
ouvertes de lecture (ORF1 et ORF2) et une deuxième région non
traduite 3' de 206 nucléotides qui se termine par une queue poly A. La
région UTR 5' joue le rôle de promoteur interne qui contrôle
la transcription de l'élément L1 par l'ARN
polymérase II (figure 2). ORF1 code pour une protéine de 40 KDa
qui se lie à l'ARN de L1 pour former une
ribonucléoprotéine cytoplasmique (RNP) et le domaine LZ a
probablement un rôle dans la dimérisation de cette
protéine. Plusieurs analyses ont montré qu'ORF1 contribue dans la
rétrotransposition mais son rôle exact n'est pas encore clair. Des
études suggèrent qu'elle a une fonction de chaperonne des acides
nucléiques pour leurs permettre des réarrangements stables. Alors
qu'ORF2 code pour une protéine de 150 kDa qui possède des
activités endonucléase et rétrotranscriptase qui va couper
l'ADN de l'hôte et faire une transcription inverse de l'ARN de
L1 en ADN qui sera inséré dans le site de coupure de
l'ADN hôte formant ainsi un nouvel élément L1.
(Goodier et Kazazian 2005)
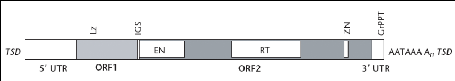
Figure 2 : Structure complète
d'un élément LINE humain (UTR : untranslated region, TSD
: variable length target site duplication; EN : endonuclease
domain; RT: reverse transcriptase domain ; Lz . a putative
leucine zipper domain ; IGS : intergenic spacer region ZN :
putative zinc knuckle motif ; GrPPT : guanosine-rich polypurine
tract) (Goodier et Kazazian 2005)
La famille de L1 peut être divisé en
deux sous-familles : les Ta (transcribed, subset a) et pre-Ta qu'on peut les
distinguer par diagnostic des trinucléotides de leur région UTR
5'. La sous-famille Ta comporte le plus grand nombre d'éléments
actifs puisque 13 des 14 événements de rétrotransposition
récemment découvert et qui sont responsable de maladies chez
l'homme sont dus à des insertions de Ta. La dernière insertion
des quatorze est due à un pre-Ta. Ce qui montre une grande
activité de cette sous-famille. (Goodier et Kazazian 2005)
Les études de Kazazian et Moran en 1998 ont
montré que le génome humain diploïde comporte
entre 80
à 100 éléments L1 actifs. Par contre le
génome de la souris compte des milliers
d'éléments
L1 actifs et qui sont subdivisés en plusieurs sous familles.
Ceci est démontré par le
pourcentage élevé de mutation chez la souris (2.5%)
par rapport à l'homme (0.07%). (Kazazian and Moran 1998)
Les éléments L1 sont rencontrés
dans tous les chromosomes mais préférentiellement sur le
chromosome X où ils représentent 26% de sa composition. Les
analyses cytogénétiques ont montré que les
séquences L1 atteignent une grande densité au niveau de
l'hétérochromatine. Des études supposent un rôle
présumé des L1 dans l'inactivation du chromosome X.
(Goodier et Kazazian 2005)
| 

