La maladie de Basedow: rôle du système immunitaire( Télécharger le fichier original )par Karim CHAJAI Université Abdelmalek Essaà¯di ; faculté des sciences Tétouan - Licence d'études fondamentales en sciences de la vie biologie cellulaire et moléculaire 2013 |

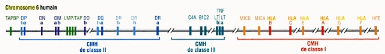
PréambuleLes maladies auto-immunes constituent la troisième cause de mortalité dans le monde après les cancers et les maladies cardiovasculaires ; ce sont la conséquence d'une réponse inappropriée des cellules du système immunitaire vis-à-vis des constituants propres de l'organisme : ces derniers sont attaqués comme s'il s'agissait de substances étrangères. Les maladies auto-immunes, telles que les dysthyroïdies auto-immunes, résultent d'une combinaison de plusieurs phénomènes immunologiques qui conduisent, au final, à la rupture de la tolérance périphérique aux antigènes du «soi» et donc à une réponse auto-immune. Ce modeste travail de mémoire sera consacré à un type particulier de dysthyroïdies auto-immunes, la maladie de Basedow ou maladie de graves. Il s'agit d'une cause principale de l'hyperthyroïdie où plusieurs facteurs, tant génétiques qu'environnementaux, contribuent au développement de la maladie et à la persistance d'auto-anticorps stimulants dirigés contre la thyroïde. Ce travail présentera, dans un premier chapitre, une revue générale sur le phénomène d'auto-immunité, puis, par un second chapitre, il apportera des généralités sur le fonctionnement normale de la glande touchée par la maladie, la glande thyroïde, pour en fin, grâce à un troisième chapitre, seront discutés certains facteurs prédisposant à l'apparition de la maladie de Basedow, les principaux symptômes, le diagnostic et les approches thérapeutiques disponibles contre cette maladie. INTRODUCTIONDans l'environnement qui nous entoure (air, eau, terre, aliments, etc.), il existe de nombreux organismes pathogènes microscopiques qui sont en contacte permanent avec notre corps, et qui représentent un véritable danger pour notre santé puisqu'ils sont souvent les causes directes de maladies. La seule chose qui nous protège de leurs dangers et qui nous permet de rester souvent en bonne santé c'est l'immunité. L'immunité est donc définie comme la résistance aux maladies, et plus spécifiquement aux maladies infectieuses. L'ensemble des cellules, des tissus et des molécules qui concourent à opposer une résistance aux infections est appelé système immunitaire (SI), et la réaction coordonnée de ces cellules et molécules contre les germes pathogènes porte le nom de réponses immunitaires ( Abbas et Lichtman, 2008). Le SI est un système diffus constitué de nombreuses structures discrètes, comprenant la moelle osseuse, le thymus, la rate, et les ganglions lymphatiques, ainsi que de nombreuses sortes de cellules sanguines circulant librement dans le sang, appelées lymphocytes : Ce sont les petites cellules rondes que l'on appelle « globules blancs ». Les cellules du SI sont produites dans la moelle osseuse et certaines reçoivent une éducation supplémentaire dans le thymus où ils terminent leur maturation, notamment les lymphocytes T (LT). Elles se déploient alors pour aller occuper l'ensemble du corps, à l'exception du cerveau, (Rensberger, 1999) prêtes à commencer leurs fonctions protectrices. La fonction physiologique du SI est de prévenir et d'éradiquer les infections déclarées ( Abbas et Lichtman, 2008) tout en répondant de façon appropriée à un nombre considérable d'agents distincts et potentiellement pathogènes qui pénètrent et envahissent l'organisme humain ( Boutammina, 2012). Le système de défense de l'organisme se compose d'une immunité innée ou naturelle, responsable de la protection initiale contre les infections (dont les éléments participants sont une surface épithéliale ou barrière, des neutrophiles et des macrophages, des cellules Natural killer, un certain nombre de protéines incluant des cytokines et des composants du système du complément ( Kierszenbaum, 2006), et d'une immunité adaptative ou acquise qui se développe plus lentement et met en oeuvre une défense tardive et plus efficace contre les infections. Cette dernière se subdivise en une immunité humorale, dans laquelle des anticorps (Ac) fabriqués par les lymphocytes B (LB) neutralisent et éliminent les microbes et les toxines extracellulaires, et en une immunité cellulaire, dans laquelle des LT éliminent les microbes intracellulaires. Ces cellules de l'immunité acquise, c'est-à-dire les lymphocytes, expriment des récepteurs qui reconnaissent, de manière spécifique, différentes substances produites par les microbes, ainsi que des molécules non infectieuses : Ces substances sont appelées antigènes (Ag). Tandis que les mécanismes de l'immunité naturelle reconnaissent des structures partagées par plusieurs classes de microbes, qui sont appelés PAMPs (Pathogen-associated molecular patterns). Le terme d'immunité innée fait référence au fait que ce type de défense développé par l'hôte est toujours présent chez les individus sains, prêt à bloquer l'entrée des microbes et à éliminer rapidement ceux qui ont réussi à pénétrer dans les tissus de l'hôte. Tandis que l'immunité adaptative est le type de défense qui est stimulé par les microbes qui envahissent les tissus, c'est-à-dire qu'elle s'adapte à la présence des micro-organismes invasifs ( Abbas et Lichtman, 2008). Généralement, tous les lymphocytes sont produits dans la moelle osseuse hématopoïétique, à partir de cellules souches multipotentes. Ces lymphocytes (B ou T) sont capables de reconnaitre une molécule ou un Ag par l'intermédiaire d'un récepteur spécifique : immunoglobuline de surface ou B cell receptor (BCR) pour le LB et T cell receptor (TCR) pour le LT. Ces récepteurs sont produits par l'association aléatoire de gènes et génèrent une capacité de reconnaissance phénoménale. Les LB peuvent reconnaitre l'Ag sans que celui-ci ne soit modifié. Les LT, en revanche, ne reconnaissent l'Ag qu'une fois dégradé et présenté dans une molécule du complexe majeur d'histocompatibilité (CMH). Ce complexe tire son nom des réactions intenses qu'il suscite lors de la transplantation d'organes incompatibles. En présentant des peptides antigéniques aux LT, ces molécules de CMH, qui chez l'Homme s'appelle antigènes leucocytaires humains (HLA, human leucocyte antigen), jouent un rôle fondamental dans la réponse immunitaire. Il s'agit de glycoprotéines de surface réparties en deux classes, I et II, caractérisées par un important polymorphisme génétique, c'est-à-dire que chaque locus comporte de multiples allèles. En conséquence, la variabilité génétique entre individus est très grande ; à moins qu'ils n'appartiennent à une famille, ils ont le plus souvent des HLA différents. Ce complexe contient au mois 128 gènes fonctionnels parmi lesquels plus de 20% ont une fonction dans l'immunité ; c'est la région du génome humain avec la densité de gènes la plus élevée. C'est aussi la région à laquelle sont associées le plus de maladies : la plupart sinon toutes les maladies auto-immunes (MAI) sont associées à des gènes du CMH, ce qui met en évidence le rôle essentiel de ces molécules dans la sélectivité des réponses immunitaires (Hachulla et Hatron, 2006 ; Chapel et al., 2004). Les gènes du CMH sont désignés par des lettres comme montré dans la figure1. Les gènes codant pour les molécules de CMH sont traditionnellement répartis en trois classes : les gènes du CMH de classe I et II qui codent pour les molécules du CMH présentatrices d'Ag, et les gènes de classe III, un groupe de gènes assez disparates codant pour d'autres molécules immunitaires (DeFranco et al., 2009).
Figure. 1. Les gènes de CMH humain (DeFranco et al., 2009). Diagramme simplifié du CMH sur le chromosome 6 humain. (Seuls les gènes de classe III qui ont une fonction immunitaire identifié sont représentés). DP, DQ, DR, A, E : Gènes classiques de classe II HLA-A, HLA-B, HLA-C, K, D, L : Gènes de classe I DN, DM, DO, M, O : Gènes non classique de classe II MICA, MICB, HLA-E, HLA-G, HLAf, HFE, Q, T, M : gènes de molécules non classiques du CMH de classe I C4A, , Bf : gènes de molécules du complément LMP : gènes de composants du protéasome, le complexe enzymatique qui dégrade les proteines en peptides dans le cytoplasme LT : gènes de la lymphotoxine TAP : gènes du transpoorteur responsable de l'entrée des peptides du cytoplasme dans le réticulum endoplasmique TAPBP : gène de la tapasine, une moélcule chapéronne ipliquée dans le chargement des peptides CMH de classe I TNF : gènes du facteur de nécrose tumorale, une cytokine importante En fait, les molécules du CMH présentatrices d'un Ag peptidique sont parfois appelées molécules CMH classiques. Il existe aussi des molécules structuralement apparentées à ces deux classes, mais dont la fonction n'est pas la présentation de l'Ag aux cellules T : elles sont appelées molécules CMH apparentées à la classe I ou à la classe II ou molécules non classiques du CMH (DeFranco et al., 2009). Ces réactions de défense immunitaire sont basées sur la capacité du SI à distinguer le « soi » du « non soi » et, donc sur la capacité à préserver l'individualité et l'intégrité de l'organisme. Le « soi » peut être défini comme l'ensemble des tissus, cellules et molécules faisant partie intégrante de l'organisme, et qui sont codés par le génome. Le « non soi » est constitué par tout le reste (Schaechter et al., 1999), et forme, d'après les auteurs, les micro-organismes (bactéries, virus, parasites et champignons), les toxines bactériennes, différents poisons, les éléments constituant les cellules tumorales, les greffons et les transplants ( Boutammina, 2012 ). Cette capacité de distinction est, en fait, un apprentissage par divers mécanismes tout au long du processus de maturation des lymphocytes. Les lymphocytes sont exposés aux Ag du "soi" dans un but d'éliminer tous les lymphocytes qui les reconnaîtront et de laisser passer ceux qui n'en reconnaitraient pas lors de leur dernier stade de développement (DeFranco et al., 2009 ; Chapel et al., 2004). Dans des conditions normales, ces réactions immunitaires de défense suffisent le plus souvent à protéger notre corps. Malheureusement, elles ne sont plus parfois capables d'assurer ces fonctions protectrices correctement. Ainsi, il résulte de ce fait des maladies. Les anomalies de fonctionnement du SI peuvent être des causes de maladies essentiellement de deux manières : par un déficit immunitaire (réponse immunitaire insuffisante) ou par une attaque immunitaire inappropriée (réponse immunitaire excessive ou erreur de cible) qu'on rencontre dans l'allergie, les MAI et les maladies de complexes immuns dans lesquelles des Ac produits en excès endommagent des tissus normaux (Sherwood et Molotchnikoff, 2006). En raison de leur capacité à distinguer les cellules étrangères, la majorité des cellules du SI se comportent correctement la plupart du temps. Elles se retiennent "poliment" d'attaquer les protéines et les cellules dont est constitué leur propre corps. Parfois, cependant, le SI peut perdre sa capacité à distinguer le « soi » du « non-soi ». Les agents des réponses immunitaires peuvent brusquement se révéler déloyaux, échapper à tout contrôle et attaquer les propres constituants de l'organisme. Les cellules du SI peuvent, par exemple, mélanger leurs gènes et leur faire subir des mutations dont les combinaisons amèneront les cellules B et T à considérer comme étrangères des protéines parfaitement normales et fonctionnelles de leur propre corps. Les attaques de ce type peuvent être fatales aux cellules du corps, les tuant aussi facilement que le feraient des bactéries. Le résultat est ce qu'on appelle des MAI (Rensberger, 1999). Il s'agit de maladies causées par le détournement de l'immunité adaptative vers des cibles constituées par des cellules et des tissus sains de l'organisme lui-même. Au cours des MAI, les tissus qui sont attaqués (les cibles des réactions auto-immunes) et les symptômes qui se manifestent sont très variés. Dans la plupart des cas, l'incidence diffère entre hommes et femmes, celles-ci en étant plus souvent victimes. Un caractère typique de ces maladies est la présence d'Ac et de cellules T spécifiques des Ag exprimé par les tissus cibles, ces Ag sont appelés auto-antigènes (auto-Ag) et les effecteurs de l'immunité adaptative qui les reconnaissent sont appelés auto-anticorps (auto-Ac) et cellules T auto-immunes (Parham, 2003). Les SI renégats ne sont malheureusement pas rares. Ils provoquent plusieurs maladies relativement courantes. Ceci comprend la sclérose en plaques (le SI attaque les cellules spéciales qui forment un revêtement isolant autour des nerfs, provoquant un « court-circuit » des nerfs, exactement comme des fils électriques ayant perdu leur isolation), l'arthrite rhumatoïde (le SI attaque les articulations, détruit leurs tissus et provoque une inflammation douloureuse ainsi qu'un gonflement), la myasthénie grave (le SI attaque les récepteurs des cellules musculaires qui reçoivent des signaux chimiques du système nerveux, laissant les muscles paralysés parce qu'ils ne peuvent plus recevoir le signal pour se mouvoir), et le diabète de types I (le SI attaque des cellules particulières à l'intérieur du pancréas, dont le travail consiste à fabriquer de l'insuline). Dans toutes ces maladies et dans bien d'autres moins connues, le SI n'est plus capable de distinguer un Ag étranger d'un élément qui appartient à son propre corps. Les cellules immunitaires attaquent les cellules natives aussi vigoureusement que si elles étaient des ennemies mortelles de leur corps (Rensberger, 1999). Les experts ont constaté que tous les organes endocriniens et pratiquement toutes les cellules endocrines pouvaient être victimes de cette auto-agression (Chapel et al., 2004). Les réactions auto-immunes peuvent alors être dirigées contre des cellules endocrines, leurs récepteurs, les hormones ou les récepteurs de celles-ci sur des cellules cibles. Il existe plusieurs mécanismes d'agression auto-immune, autrement-dit plus d'un mécanisme pouvant intervenir dans une maladie donnée. Les cellules T et les Ac semblent opérer en parallèle dans la pathogénie de la maladie endocrine auto-immune. Grosso modo, on peut dire que les cellules T (tant CD4+ que CD8+) sont responsables de la destruction glandulaire et que les Ac perturbent la fonction physiologique de la glande affectée (Chapel et al., 2004). La thyroïde est parmi les glandes endocrines considérées comme cibles potentielles des attaques erronées du SI. Elle intervient par ses hormones, la tri-iodothyronine (T3) et la thyroxine (T4) sécrétées sous l'influence de la TSH (thyroid-stimulating hormone) antéhypophysaire, dans la régulation du métabolisme des lipides, des protides et des glucides. Ces hormones régissent notamment l'oxydation du glucose par les cellules et sa conversion en énergie chimique. Puisque toutes les cellules de l'organisme dépendent de l'apport continu d'énergie, elles sont donc toutes des cibles pour les hormones thyroïdiennes (HT). Ces dernières, par leurs actions métaboliques, participent également à la thermorégulation (effet thermogène ou calorigène), ainsi qu'au métabolisme hydrique et minéral. Elles sont essentielles à la croissance et au développement du squelette, des muscles, des organes génitaux et du système nerveux. Elles agissent sur les tissus pour augmenter la réactivité nerveuse, mais aussi la fréquence cardiaque et la pression artérielle. Toutes déficiences ou excès en hormones thyroïdiennes sont donc extrêmement perturbateurs pour l'organisme (Cornet, 2009). Les cellules thyroïdiennes expriment de nombreux Ag, pouvant servir de cibles aux auto-Ac : thyroperoxydase (enzyme clé de la synthèse des HT, TPO), thyroglobuline (précurseur des HT, Tg) et récepteur de la TSH (R-TSH), qui rendent, malheureusement, la thyroïde susceptible d'être attaquée par des lésions auto-immunes par un mécanisme mal élucidé où les immunités à médiation humorale et cellulaire interviendraient toutes les deux amenant à des anomalies du fonctionnement, ou même, à la destruction de cette glande (Perlumuter et Hernandez 2002). Les R-TSH constituent l'interface indispensable entre le thyrocyte (cellule endocrine de la thyroïde) et la TSH (hormone hypophysaire régulant la fonction et la croissance des cellules thyroïdiennes) (Kubab et Kubab 2011). Il arrive que des Ac se trompent et attaquent ces récepteurs, perturbant leur déroulement normal, ce sont les Ac anti-R-TSH (TRAK). Ces TRAK se combinent au R-TSH et miment les effets de la TSH (Weil et Batteux, 2003) provoquant une stimulation prolongée des cellules de la thyroïde qui aboutit à l'hyperthyroïdisme (augmentation de la teneur du sang en HT) (Karp 2010). Ainsi, le métabolisme est sur stimulé et passe en surrégime. Du coup, le coeur bat trop vite, les muscles sont hypertoniques, le système nerveux est en état d'alerte permanente, etc (Nys, 2012). De telles réactions sont rencontrées dans une maladie dite maladie de Basedow relative au médecin allemand Carl Adolph von Basedow qui, en 1840, fut le premier à décrire une hyperfonction de la thyroïde, désormais connue sous le nom de maladie de Basedow. Il s'agit essentiellement d'une hyperthyroïdie d'origine auto-immune. Par ailleurs, Il est possible de rencontrer l'état contraire, une hypothyroïdie, suite à une destruction totale de la glande. Qu'est-ce que c'est que l'auto-immunité ? Pourquoi et comment intervient-elle dans l'apparition de l'hyperthyroïdie ? Comment une hyperthyroïdie perturbe tout le métabolisme de l'organisme ? Y a-t-il une solution pour contrer cette mésaventure ? |
|