Production de domaines recombinants PRODH en vue de l'analyse structurale & Caractérisation de la région 51-160 de la protéine KIN17 humaine par RMN et Modélisation Moléculaire( Télécharger le fichier original )par Ludovic Carlier Université de Rouen - Doctorat de biologie structurale 2006 |

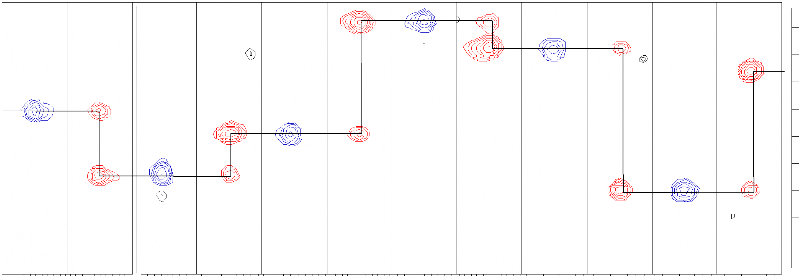
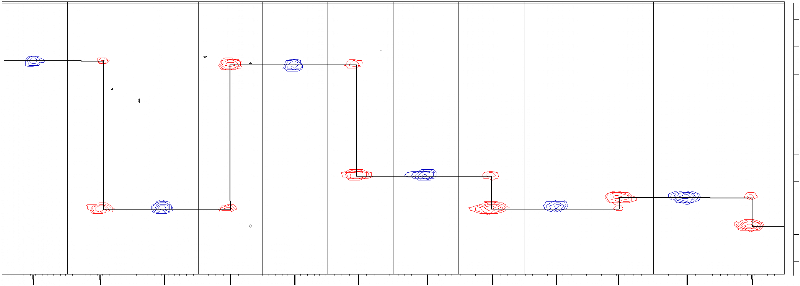
CHAPITRE 4Caractérisation structurale du domaine K2par RMN et Modélisation Moléculaire1) Détermination de la structure du domaine K2 de KIN17 humaine1.1) Attribution des raies de résonance L'attribution des raies de résonance des 111 acides aminés du domaine K2 a été réalisée selon la stratégie définie dans le chapitre précédent. Les différentes expériences 3D hétéronucléaires utilisées pour déterminer le déplacement chimique de chaque noyau, et recueillir les contraintes expérimentales, ont été enregistrées à 30°C et à pH 6.0 sur un spectromètre 600 MHz équipé d'une cryosonde triple résonance 1H, 15N, 13C. Un échantillon unique de protéine doublement marquée 15N / 13C concentrée à 0.7 mM a suffi pour mener à bien l'attribution séquentielle du squelette peptidique. La combinaison des expériences HNCO, (HCA)CO(CAN)NH, HNCA, CBCANH, et CBCA(CO)NH a conduit à l'identification des valeurs de déplacement chimique des carbones 13CO, 13Cá, et 13Câ des résidus i et i-1 relatifs à chaque groupement amide. Il est à noter que l'expérience (HCA)CO(CAN)NH, plus sensible que HN(CA)CO, a été préférée pour obtenir les corrélations des 13CO intra- et inter-résidus. De plus, l'expérience HN(CO)CA, qui a également été enregistrée, n'a pu être exploitée en raison d'un problème d'artéfacts. L'interprétation de ces 5 expériences par paire a permis dans un premier temps de connecter les résidus deux à deux, puis de reconstituer l'ensemble de la séquence peptidique à partir des acides aminés facilement identifiables tels que les glycines, thréonines, sérines, alanines, et prolines. La Figure 4.1 présente un exemple d'attribution séquentielle de la région F23-L28. L'attribution de la chaîne principale a finalement été complétée par l'analyse des spectres HNHA et HBHA(CO)NH. L'optimisation préalable des conditions d'analyse a permis d'obtenir des spectres de qualité qui contiennent une quantité d'information satisfaisante. Dès lors, la bonne dispersion des pics sur le spectre HSQC 15N-1H a facilité l'attribution de la totalité des résonances 1HN, 1Há, 15NH, 13CO, et 13Cá du domaine K2 à l'exception des résidus G1, Q2, et R53 dont les déplacements chimiques du groupement amide sont manquants. L'absence de corrélation 15N-1H de résidu appartenant à une extrémité flexible N- ou C-terminale est un phénomène classique qui s'explique par l'échange rapide du proton amide exposé au solvant avec les protons de l'eau. Aussi, en anticipant sur la résolution de la structure tridimensionnelle, F23 F23 R24 R24 N25 N25 D26 D26 F27 F27 L28 L28 176.5 177.2
A) 178.0 178.8 179.5 ppm 13CO F23 F23 R24 R24 N25 N25 D26 D26 F27 F27 L28 L28 B) 13Cá E22 13Cá F23 13Cá R24 13Cá
N25 13Cá D26 13Cá F27 56.0 58.0 60.0 62.0 64.0 ppm F23 F23 R24 R24 N25 N25 D26 D26 F27 F27 L28 L28 26.0 13Câ E22 C) 13Câ R24 30.0 34.0 13Câ 13Câ F23 13Câ
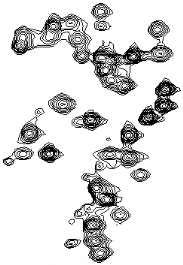
N25 13Câ D26 13Câ F27 42.0 8.61 8.61 8.48 8.48 8.72 8.72 8.97 8.97 8.81 8.81 8.43 8.43 1HN ppm Figure 4.1 : Attribution des fréquences 13CO, 13Cá, et 13Câ de la région F23-L28 du domaine K2. Les bandes sont extraites des plans 15N des spectres, A) HNCO (en bleu) et (HCA)CO(CAN)NH (en rouge), B) CBCA(CO)NH (en bleu) et HNCA (en rouge), et C) CBCA(CO)NH (en bleu) et CBCANH (en rouge) au déplacement chimique 1HN des résidus correspondants. Le chemin d'attribution est indiqué par des lignes horizontales fléchées. l'échange rapide du proton amide R53 peut être expliqué par son appartenance à un résidu situé dans une boucle, et à sa forte exposition au solvant. Les fréquences de résonance des noyaux 1H et 13C des chaînes latérales ont été déterminées à partir des spectres HCCH-TOCSY, HCCH-COSY, et 13C-NOESY-HSQC. Un échantillon unique de protéine doublement marquée 15N / 13C concentrée à 0.7 mM, préalablement lyophilisée, puis solubilisée dans du D2O (99.9 %) a suffi pour enregistrer l'ensemble de ces expériences sur les régions aliphatiques et aromatiques. La totalité des noyaux des chaînes latérales aliphatiques des 111 acides aminés de K2 a été attribuée. En ce qui concerne les chaînes aromatiques, toutes les fréquences de résonance des résidus histidine, tyrosine, et tryptophane ont été identifiées à l'exception des proton et carbone Z2 du résidu W63. Le bilan de l'attribution est plus mitigé pour les phénylalanines, et seules 3 des 6 chaînes latérales ont été totalement attribuées (F19, F23, et F15). Par ailleurs, l'expérience 15N-TOCSY-HSQC a été enregistrée sur un échantillon uniformément marqué 15N afin de vérifier l'attribution. Cependant, compte tenu de la faible efficacité du transfert TOCSY 1H notamment due à la taille de la protéine K2, moins de la moitié des valeurs de déplacement chimique aliphatique 1H a pu être contrôlée. La dernière étape de l'attribution du domaine K2 a consisté à enregistrer le spectre 15N-NOESY-HSQC sur l'échantillon 15N, ce qui a permis d'attribuer la totalité des 13 groupements NH2 de résidus asparagine et glutamine à l'exception de N42. L'identification des fréquences de résonance du domaine K2 de la protéine KIN17 humaine a fait l'objet d'une note d'attribution soumise et acceptée dans la revue Journal of Biomolecular NMR (Carlier et al., 2006). Cet article, présenté dans les pages suivantes, est accompagné d'un supplément technique (Supplemental Material) disponible sur le site de J.Biomol.NMR. à l'URL suivante : http://dx.dio.org/10.1007/10858-006-0013-y. Journal of Biomolecular NMR (2006) (c) Springer 2006 Letter to the Editor NMR assignment of region 51-160 of human KIN17, a DNA and RNA-binding protein DOI 10.1007/s10858-006-0013-y KIN17 is a 45 kDa nuclear protein which is remarkably conserved in eukaryotes, ubiquitously expressed in mammals, and forms intra-nuclear foci in proliferating cells. Major features of KIN17 are its ability to bind DNA and RNA and to be up-regulated in response to UV and irradiation, suggesting a role in DNA replication, the DNA damage response or RNA processing (Pinon-Lataillade et al., 2004; Miccoli et al., 2005). Human KIN17 comprises an N-terminal zinc finger (27-50) and a C-terminal KOW motif (335-373). Here we report the 1H, 15N, 13C assignments of the region 51-160 of human KIN17 as a preliminary step toward obtaining the atomic structure of this region and elucidating its biological function. Residues corresponding to the KIN17 domain were all identified: 99% of the backbone chemical shifts, and for the side chains, all the aliphatic and 88% of the aromatic 1H-13C nuclei were assigned. BMRB deposit with accession number 6938. References: Miccoli et al. (2005) Mol. Cell. Biol., 25, 3814-3830; Pinon-Lataillade et al. (2004) J. Cell. Sci., 117, 3691-3702. Ludovic Carliera, Albane le Maireb, Sandrine Braudb, Cedric Massonb, Muriel Gondryb, Sophie Zinn-Justina, Laure Guilhaudisa, Isabelle Milazzoa, Daniel Davousta, Bernard Gilquinb & Joël Couprieb,* aEquipe de Chimie Organique et Biologie Structurale, CNRS UMR 6014, IFRMP 23, Université de Rouen, France; bDépartement d'Ingénierie et d'Etude des Protéines, CEA Saclay, 91191, Gif-sur-Yvette, France *To whom correspondence should be addressed. E-mail: jo el.couprie@cea.fr Supplementary material is available in electronic format at http://dx.dio.org/10.1007/10858-006-0013-y. Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire Letter to the Editor (Supplemental Material) NMR assignment of region 51-160 of human KIN17, a DNA and RNA-binding protein Ludovic Carliera, Albane le Maireb, Sandrine Braudb, Cédric Massonb, Muriel Gondryb, Sophie Zinn-Justinb, Laure Guilhaudisa, Isabelle Milazzoa, Daniel Davousta, Bernard Gilquinb and Joël Couprieb,* a Equipe de Chimie Organique et Biologie Structurale, IFRMP 23, CNRS UMR 6014, Université de Rouen b Département d'Ingénierie et d'Etude des Protéines, CEA Saclay, 91191 Gif-sur-Yvette, France. 187 * To whom correspondence should be addressed : joel.couprie@cea.fr Keywords: NMR assignments, KIN17, DNA-binding, RNA-binding, nuclear metabolism Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire Biological context KIN17 is a 45 kDa nuclear protein conserved in eukaryotes and initially identified on the basis of its cross-reactivity with antibodies raised against bacterial RecA, a recombination-repair enzyme (Kannouche et al., 2000). In mammals, the KIN17 protein is ubiquitously expressed. It forms intranuclear foci in proliferating cells and is up-regulated after UV or ã irradiation (Biard et al., 2002; Kannouche et al., 1998). The other features of KIN17 are its ability to bind DNA and RNA in vitro and in vivo (Biard et al., 2002; Pinon- Lataillade et al., 2004), to associate with multiprotein DNA replication complexes (Miccoli et al., 2005), and to complement the functions of the bacterial transcriptional factor H-NS (Timchenko et al., 1996), suggesting a role in nuclear metabolism. Human KIN17 is a modular protein comprising four motifs: a zinc finger (28-50) at the N-terminus, a core domain homologous to RecA protein (163-201), a nuclear localization signal (240-257), and a KOW motif (335-373) found in several bacterial transcription factors (Kannouche et al., 2000; Ponting et al., 2002). On the basis of sequence alignment and secondary structure prediction, we identified a globular region located between the zinc finger and the core domain. We here report the 1H, 15N, 13C assignments of the region 51-160 as a preliminary step toward obtaining the atomic structure of this KIN17 domain and elucidating its biological function. Methods and experiments 188 The gene encoding the region 51-160 of human KIN17 was amplified by polymerase chain reaction (PCR) and cloned into several expression vectors using the Gateway system (Invitrogen). A protein expression screening using different strains and fusion partners was performed (Braud et al., in press) and highest levels of expression and solubility were obtained when transforming E. coli Rosetta(DE3) strain with plasmid pEXP-TH5 (Invitrogen). This vector encodes a 6xHis tag, the Z-domain from Staphylococcal Protein A (ZZ), a TEV protease (Tobacco Etch Virus) cleavage site, and the KIN17 51-160 domain. Protein expression was induced by the addition of 1 mM of isopropyl-1-thio-â-D- galactopyranoside (IPTG) to the bacterial culture at a cell density of 1.2 absorbance units measured at 595 nm. The bacterial culture was further grown for 14 hours at 20°C. The fusion protein was purified using IgG Sepharose Fast Flow (Amersham) in order Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire to separate the fusion protein from other bacterial proteins and to remove the (His)6-ZZ tag after an on-column cleavage using recombinant (His)6-TEV protease. A second step of purification was performed using Ni-NTA column (Qiagen) to remove (His)6-TEV protease and residual (His)6-ZZ tag. Uniformly labelled 15N protein was produced in minimal medium M9 containing 1 g.L-1 of (15NH4)2SO4 (Boehringer) as the sole nitrogen source. Uniformly labelled 13C/ 15N protein was produced in a rich medium prepared from uniformly labelled 13C/15N Spirulina maxima cyanobacteria. NMR samples containing about 0.7 mM protein were prepared in 50 mM phosphate buffer pH 6.0 containing 150 mM NaCl, 1 mM EDTA, a protease inhibitor cocktail (SIGMA), 1 mM TCEP, and 1 mM NaN3 in either 90% H2O/10% D2O or in 100% D2O. 3-(trimethylsilyl)[2,2,3,3-2H4] propionate (TSP) was added as an internal 1H chemical shift reference. 13C and 15N chemical shifts were referenced indirectly to TSP, using the absolute frequency ratios (Wishart et al., 1995). NMR experiments were performed at 303 K on Bruker AVANCE DMX-600 or Varian INOVA-800 spectrometers equipped with triple-resonance (1H, 15N, 13C) cryoprobes. Spectra used for sequential backbone assignment were as follows: 2D 15N-1H HSQC, 3D HNCO, 3D HNCA, 3D CBCA(CO)NH, 3D CBCANH, 3D HNHA, 3D (HCA)CO(CAN)NH experiments. Aliphatic and aromatic 1H and 13C side chain assignments were obtained using 3D HBHA(CO)NH, 3D HCCH-TOCSY, 3D HCCH-COSY, and 3D 13C-HSQC NOESY experiments. All NMR data were processed with NMRPipe software (Delaglio et al., 1995) and analysed with Felix software (Molecular Simulations). 189 Extent of assignments and data deposition For cloning strategy reasons, this KIN17 domain contains the region 51-160 of human KIN17 plus an additional glycine residue at the N terminus. Figure 1 shows the 15N-1H HSQC spectrum of the 111 residues protein numbered from 1 to 111. All the amide 1H and 15N backbone resonances were assigned except for N-terminal amino acids G1 and Q2, and R53 whose NH is unobservable presumably due to loop mobility or chemical exchange since this residue is located between secondary structure elements. Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire Compared to the expected number of resonances, nine additional weakened cross- peaks were observed in the 15N-1H HSQC spectrum. However, backbone and side chain assignments, as well as 3D 15N HSQC-NOESY patterns revealed that 6 spin systems split in two (R3, Q4, L5, L6, A8, S9). Those residues are located before the proline P12 and we suppose that the minor form is due to a partial isomerisation of P12. The observation of a characteristic strong NOE between Hä of P12 and Há of N11 indicated that the predominant form of this proline is in a trans conformation. Finally, all residues corresponding to the region 51-160 of KIN17 were identified: the backbone assignment is complete for 1Há, 13Cá and 13CO, and it reaches 98% for 1HN and 15N nuclei. All the aliphatic side chain resonances were assigned and 88% of the aromatic 1H and 13C resonances were identified, including all Tyr and His aromatic rings, 3 Phe out of 6, and 2 Trp out of 3. Assignments of NH2 side chain resonances of asparagines and glutamines were also completed except for N42. The 1H, 13C and 15N chemical shifts have been deposited in the BioMagResBank (http://www.bmrb.wisc.edu) under accession number 6938. Acknowledgement The Bruker DMX-600 spectrometer was supported by grants of the Conseil Regional de Haute-Normandie (France). We are grateful to Dr Adrien Favier who kindly recorded 3D HSQC NOESY experiments on Varian INOVA-800 spectrometer at I.B.S Jean-Pierre Ebel (Grenoble, France), and to Marie Courçon and Mireille Moutiez for the protein expression screening. We also thank the Centre de Ressources Informatiques de Haute-Normandie (France) for NMR software facilities. References Braud, S., Moutiez, M., Belin, P., Abello, N., Drevet, P., Zinn-Justin, S., Courçon, M., Masson, C., Dassa, J., Charbonnier, J.B., Boulain, J.C., Ménez, A., Genet, R., Gondry, M. (2005). J. Proteome Res., In press. Biard, DS., Miccoli, L., Despras, E., Frobert, Y., Creminon, C. and Angulo, JF. (2002) 190 J. Biol.Chem., 277(21), 19156-19165. Delaglio, F., Grzesiek, S., Vuister, G. W., Zhu, G., Pfeifer J. and Bax, A. (1995) J. Biomol. NMR., 6, 277-293. Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire Kannouche, P., Mauffrey, P., Pinon-Lataillade, G., Mattei, MG., Sarasin, A., Daya-Grosjean, L. and Angulo, JF. (2000) Carcinogenesis, 21(9), 1701-1710. Kannouche, P., Pinon-Lataillade, G., Tissier, A., Chevalier-Lagente, O., Sarasin, A., Mezzina, M. and Angulo, JF. (1998) Carcinogenesis, 19(5), 781-789. Miccoli, , L., Frouin, I., Novac, O., Di Paola, D., Harper, F., Zannis-Hadjopoulos, M., Maga, G., Biard, D.S.F., Angulo, J.F. (2005). Mol. Cell. Biol., 25(9), 3814-3830. Pinon-Lataillade, G., Masson, C., Bernardino-Sgherri, J., Henriot, V., Mauffrey, P., Frobert, Y., Araneda, S. and Angulo, JF. (2004) J. Cell. Sci., 117(16), 3691-3702 Ponting, CP. (2002) Nucleic Acids Res., 30(17), 3643-3652. Timchenko, T., Bailone, A. and Devoret, R. (1996) EMBO J., 15, 3986-3992. Wishart, DS., Bigam, CG., Yao, J., Abildgaard, F., Dyson, HJ., Oldfield, E., Markley, JL. and Sykes, BD. (1995) J. Biol. NMR, 6, 135-140 R33 N11 E22 L73 K71 G74 T65 Q13 Y46 L31 E29 R102 E105 E107 T84 Q4 D17 T67 E10 Q91 Y18 L6 Q103 L104 M16 F15 Q4 R101 K109 K38 K108 L106 T61 G87 G77 D96 E83 R39 Y49 L7 R95 K110 L5 W63 A8 I90 E21 N59 N47 T37 R34 F35 I50 G36 S9 K86 E76 T70 Q62 C79 S51 E54 N25 S20 R24
L28 F19 R32 T99 L78 H55 V45 E64 D26 E98 F23 R3 F69 L30 I100 A60 E48 I44 H52 Y89 N43 F27 W88 L66 D82 V81 I56 K80 M58 V40 R75 D94 D68 W72 I93 N42 H57 191 H41 K111 W63 å W88 å Y92 W72 å Figure 1: 2D 15N-1H HSQC spectrum of the region 51-160 of human KIN17. The assignments are indicated with the one-letter amino acid code. NH2 side chain resonances of asparagines and glutamine are connected by purple horizontal lines. Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire 1.2) Détermination de la topologie du domaine K2 La première étape de la caractérisation structurale du domaine K2 a consisté à déterminer les éléments de structure secondaire à partir de paramètres structuraux et dynamiques RMN afin d'établir la topologie de la protéine. 1.2.1) Analyse du nOe hétéronucléaire 1H-15N Nous avons dans un premier temps réalisé une expérience de mesure des nOe hétéronucléaires 1H-15N dans le but d'identifier les résidus non structurés des extrémités N- et C-terminales. Les spectres ont été enregistrés à 600 MHz sur un échantillon uniformément marqué 15N concentré à 0.8 mM. 1 0.8 0.6 0.4 0.2 0 ** * * * * * * -0.2 10 20 30 40 50 60 70 80 90 100 -0.4 -0.6 -0.8 -1 Figure 4.2 : nOe hétéronucléaires 1H-15N du domaine K2 de la protéine humaine KIN17. Les astérisques correspondent aux résidus dont la valeur n'a pas été déterminée ou aux résidus proline. Les faibles valeurs de nOe hétéronucléaire (<0.25) observées au niveau des extrémités N- et C-terminales mettent en évidence que les régions R3-N11 et K108-K111 sont flexibles et déstructurées (Figure 4.2). L'échange rapide du proton amide de Q2 avec l'eau n'ayant pas permis de mesurer sa valeur de nOe 1H-15N, et le résidu 12 étant une proline, il semble raisonnable de conclure que les segments G1-P12 et K108-K111 sont déstructurés. 192 Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire 1.2.2) Analyse des paramètres structuraux classiques La caractérisation des éléments de structure secondaire a été initiée en calculant les indices de déplacement chimique CSD à partir des valeurs de déplacements chimiques Há et Cá, et des valeurs de référence correspondantes dans la BioMagResBank. La figure 4.3 montre le diagramme des indices ?äCá et ?äHá du domaine K2. La majorité des résidus présente une valeur positive de ?äCá conjuguée à une valeur négative de ?äHá, ce qui suggère une structuration hélicoïdale importante. Pour être significatives, les valeurs de ?äCá et ?äHá doivent atteindre respectivement, 1, et 0.1 ppm. Sur la base de ces valeurs seuils, 4 hélices sont clairement identifiables au niveau des régions D17-R33, H41-S51, L66-R75, et P97- E107. A contrario, la région C79-Q91 se caractérise plutôt par des valeurs négatives de ?äCá conjuguées à des valeurs positives de ?äHá, ce qui indique une conformation en feuillet â ou étendue. Cependant, le signe des indices CSD dans cette région est irrégulier, et l'amplitude des index ?äCá négatifs est relativement faible. Afin d'approfondir la caractérisation des structures secondaires, nous avons entrepris une analyse partielle du spectre 15N-NOESY-HSQC enregistré à 600 MHz sur un échantillon uniformément marqué 15N concentré à 0.8 mM. Selon la stratégie définie dans le chapitre 2, nous nous sommes contentés de collecter 3 types de nOe non ambiguës facilement identifiables. Il s'agit des corrélations caractéristiques de type dáN(i, i+1), dáN(i, i+2), et dáN(i, i+3). Nous avons également estimé les constantes de couplage 3JHN-Há à partir des expériences 3D HNHA et 2D HMQC_J enregistrées sur l'échantillon 15N. L'analyse combinée des nOe caractéristiques et des constantes de couplage 3JHN-Há (Figure 4.4) permet de préciser la nature des éléments de structure secondaire mis en évidence précédemment : L'observation de contacts dáN(i, i+3) consécutifs associés à de faibles intensités de nOe dáN(i, i+1), et à une absence d'effets dáN(i, i+2), caractérise une structuration des 4 régions D17-L31, N42-S51, L66-G77, et P97-Q103 en hélice á. En ce qui concerne l'hélice C-terminale, la forte similarité des valeurs de déplacement chimique Há dans la région L104- K110 ne permet pas de relever d'effets supplémentaires non ambiguës au delà du résidu 103. Les valeurs consécutives de 3JHN-Há inférieures à 5.5 Hz au niveau des régions F15-R33, H41- Y49, D68-R75, et D96-E105 confirment l'existence de ces 4 hélices. Notons toutefois que 3 des valeurs de constante 3JHN-Há du segment H41-Y49 (I44, V45, et E48) se situent entre 5.5 et 6 Hz. 193 5 10 15 20 25 30 35 Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire 7
6 5 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110 4 3 2 1 0 -1 -2 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110 194 -3 1.2 0.9 0.6 0.3 0 -0.3 -0.6 -0.9 -1.2 -1.5 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110 -1.8 Figure 4.3 : Déplacements chimiques secondaires du domaine K2 de la protéine KIN17 humaine calculés à partir de valeurs de référence de la 194 BioMagResBank (http://www.bmrb.wisc.edu/) Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110 | | | | | | | | | | | | | | | | | | | | | | GQRQLLLASENPQQFMDYFSEEFRNDFLELLRRRFGTKRVHNNIVYNEYISHREHIHMNATQWETLTDFTKWLGREGLCKVDETPKGWYIQYIDRDPETIRRQLELEKKKK dáN(i,i+1) dáN(i,i+3) 195 dáN(i,i+2) J 3 9 HN-Há 7 [Hz] 7 6 5 4 3 2 1 0 5 3 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110
GQRQLLLASENPQQFMDYFSEEFRNDFLELLRRRFGTKRVHNNIVYNEYISHREHIHMNATQWETLTDFTKWLGREGLCKVDETPKGWYIQYIDRDPETIRRQLELEKKKK Figure 4.4 : Diagramme récapitulatif de 3 effets Overhauser caractéristiques et des constantes de couplage 3JHN-Há du domaine K2 de KIN17 humaine. L'épaisseur des traits rouges, bleus, et verts indique respectivement, l'intensité des effets à courte et moyenne distance de type dáN(i, i+1), dáN(i, i+3), et dáN(i, i+2) relevés sur le spectre NOESY-HSQC édité 15N. Les lignes horizontales violettes représentent les valeurs seuil de constante de couplage 3JHN-Há dans les structures secondaires (<5.5 Hz dans l'hélice á ou 310, >8 Hz dans le feuillet â). Les traits 195 inclinés correspondent aux résidus dont la valeur de 3JHN-Há n'a pas été déterminée. De manière générale, peu de connexions non ambiguës de type dáN(i, i+2) ont été relevées sur le spectre 15N-NOESY-HSQC. Ces effets sont caractéristiques d'une structuration en hélice 310 ou en coude. L'observation de 2 contacts consécutifs de moyenne intensité de type dáN(i, i+2) et d'un effet dáN(i, i+3) au niveau du segment M58-T61 suggère l'existence potentielle d'un tour d'hélice 310. Cette hypothèse est consolidée par une faible valeur de 3JHN-Há (<4.5 Hz), et une valeur significative d'indice ?äCá des résidus M58 et N59. La succession d'effets dáN(i, i+1) de forte intensité associés à une absence quasi-totale d'effets dáN(i, i+3) et dáN(i, i+2) au niveau des régions C79-T84 et W88-D94 confirme la présence d'une zone étendue ou structurée en feuillet â dans la région C79-D94. Les fortes valeurs de constante 3JHN-Há (>8 Hz) au niveau des résidus K80 à D82, T84, K86, Y89 à Q91, et I93 sont en accord avec ce résultat. D'autre part, le diagramme de la Figure 4.4 fait apparaître 4 fortes intensités dáN(i, i+1), un contact dáN(i, i+2), et 3 valeurs de 3JHN-Há supérieures à 8 Hz dans la région T35-N42. Ce profil laisse présager la présence d'un ou plusieurs types conformationnels : coude(s), brin â, ou zone étendue. Aussi, il est difficile de caractériser la structure secondaire de cette zone à partir de ces seules données. Les valeurs significatives d'indice ?äHá des résidus T37, R39 et V40 (> +0.2 ppm), et la forte valeur de ?äCá du résidu V40 (-1.9 ppm) supportent l'hypothèse de l'existence d'un brin â. 1.2.3) Prédiction des angles et Le déplacement chimique des noyaux 13Cá, 13Câ, 13CO, 1Há, et 15NH a été utilisé pour estimer les angles dièdres et caractéristiques des structures secondaires à l'aide du logiciel TALOS. Dans le cas du domaine K2, 68 des 111 valeurs de couple d'angles et prédites sont considérées comme fiables. Une grande partie de ces valeurs se situe dans la région spécifique aux hélices á du diagramme de Ramachandran, ce qui confirme une structuration hélicoïdale importante. Sur la base de cette prédiction, 4 hélices á sont identifiables au niveau des régions Y18-R34, N42-I50, T67-R75, et E98-L106. Le programme TALOS prédit également la présence de 3 zones étendues ou structurées en brin â au niveau des segments R39-H41, C79-T84, et W88-Y92. De plus, les valeurs d'angles dièdres prédites pour les résidus M58 et N59 sont caractéristiques d'une hélice 310, ce qui conforte l'hypothèse de la présence d'un tel motif dans cette région du domaine K2. Par ailleurs, les valeurs particulières d'angles et créditées aux résidus isolés R53 et E54, ainsi qu'aux résidus E76, P85, et K86, suggèrent une implication de ces acides aminés dans un coude régulier. 1.2.4) Identification et organisation des brins â Afin de préciser la structure secondaire des régions qui présentent une préférence conformationnelle étendue, nous avons entrepris une recherche d'effets spécifiques à longue distance de type HN-HN, HN-Há, et Há-Há sur les spectres 15N- et 13C-NOESY-HSQC. L'analyse de ces effets met en évidence l'existence d'un feuillet â anti-parallèle triple brin dans lequel le brin central G87-I93 est encadré par un second de même taille qui le précède dans la séquence (L78-T84), et par un troisième beaucoup plus court (V40-N42) (Figure 4.5). Les résultats de l'expérience de spectroscopie d'échange 1H ? 2D menée sur un échantillon 15N / 13C solubilisé dans 100% D2O sont en accord avec une telle organisation : le ralentissement constaté de la vitesse d'échange des protons amides T84, Y89, I90, Q91, et I93 suggère une implication de ces protons dans une liaison hydrogène, et confirme leur appartenance à une structure secondaire régulière (Figure 4.5). L78 C79 V81 K80 D82 E83 T84 P85 I93 Y92 Q91
I90 Y89 W88 G87 K86 V40 N42 H41 Figure 4.5 : Reconstitution de la topologie du feuillet â anti-parallèle du domaine K2 à partir de l'analyse des nOe longue distance Há-Há (traits oranges), HN-HN (traits bleus), et HN-Há (traits verts). Les lignes hachurées correspondent aux liaisons hydrogène déduites de l'analyse des vitesses d'échange des protons amides et des nOe inter-brins. 1.2.5) Conclusions : structure secondaire du domaine K2 La prise en compte de l'ensemble des informations structurales obtenues à partir des indices CSD, des constantes 3JHN-Há, des nOe 1H caractéristiques, des nOe hétéronucléaires, de l'échange des protons amides, et de la prédiction des angles et , permet finalement de caractériser la topologie des structures secondaires (Figure 4.6). Le consensus réalisé à partir de toutes ces informations met clairement en évidence l'existence de 4 hélices á (H1 à H4) et de 3 brins â (S1 à S3). De plus, comme nous l'avons suggéré dans les paragraphes précédents, plusieurs éléments sont en faveur de l'existence d'un tour d'hélice 310 dans la région M58- T61. A ce stade de l'étude structurale, la présence de ce motif demeure une simple hypothèse qui doit être confirmée par le calcul de la structure par modélisation moléculaire. Il en est de même pour l'orientation et la délimitation précise des structures secondaires, uniquement accessibles par la détermination de la structure tridimensionnelle à l'échelle atomique.
GQRQLLLASENPQQFMDYFSEEFRNDFLELLRRRFGTKRVHNNIVYNEYISHREHIHMNATQWETLTDFTKWLGREGLCKVDETPKGWYIQYIDRDPETIRRQLELEKKKK nOe 15N CSD 3JHN-Há TALOS nOe 1H Consensus
GQRQLLLASENPQQFMDYFSEEFRNDFLELLRRRFGTKRVHNNIVYNEYISHREHIHMNATQWETLTDFTKWLGREGLCKVDETPKGWYIQYIDRDPETIRRQLELEKKKK
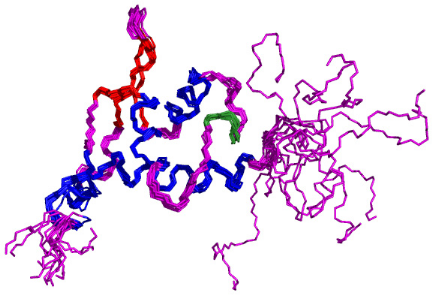
Figure 4.6 : Caractérisation de la structure secondaire du domaine K2 par RMN. Un cylindre bleu correspond à une hélice á, un cylindre vert à une hélice 310, une flèche rouge à un brin â, et un rectangle rouge à une région étendue ou à un brin â. Les traits noirs prononcés indiquent l'étendue des extrémités flexibles et déstructurées et une étoile correspond à un résidu dont la vitesse d'échange de proton amide est ralentie. 1.3) Calcul de la structure par Modélisation Moléculaire sous contraintes RMNLa structure du domaine K2 de la protéine KIN17 humaine a été calculée avec le logiciel CNS couplé au programme d'attribution automatique des nOe du Laboratoire de Structure des Protéines. Les méthodes et principes sur lesquels reposent ces logiciels sont exposés dans le chapitre 3 de cette seconde partie. 1.3.1) Préparation des contraintes expérimentales RMN Les spectres NOESY-HSQC utilisés pour déduire les contraintes de distance ont été enregistrés à l'IBS de Grenoble sur un spectromètre 800 MHz équipé d'une cryosonde triple résonance, avec un temps de mélange de 80 ms pour l'expérience éditée 15N, et de 100 ms pour l'expérience éditée 13C. Les pics de corrélation dipolaire ont été sélectionnés, intégrés, puis collectés à l'aide du logiciel Felix (Accelrys). Aucun de ces pics n'a été attribué manuellement à l'exception des 9 effets à longue distance de type HN-HN, HN-Há, et Há-Há qui définissent la topologie du feuillet â de K2. Les 136 valeurs d'angles et prédites par le logiciel TALOS ont été utilisées pour former les contraintes d'angle dièdre. Les intervalles de valeurs permises associés à ces angles ont été établis en multipliant l'écart type estimé par TALOS pour chaque prédiction par un coefficient d'une valeur de 1.5. Trois contraintes supplémentaires d'angle ont été ajoutées dans les protocoles de calcul sur la base d'une valeur de constante de couplage 3JHN-Há caractéristique des résidus F35, H55, et D96. Les intervalles de valeurs permises correspondants ont été déduits de la courbe de Karplus lissée en utilisant les coefficients de Pardi (Pardi et al., 1984). Enfin, l'expérience de spectroscopie d'échange 1H ? 2D et la caractérisation des éléments de structure secondaire ont permis de mettre en évidence l'existence de 18 liaisons hydrogène principalement localisées dans les hélices á (Figure 4.6). Ces informations ont été interprétées en contrainte de distance (dHN-O ~ 2.3 Å et dN-O ~ 3.3 Å) et introduites dans les protocoles de calcul. 1.3.2) Bilan de l'attribution automatique des pics nOe Le calcul de la structure du domaine K2 a nécessité la répétition de 21 processus itératifs de 22 cycles où chaque itération a fait l'objet d'une attribution des pics nOe et d'un calcul de 200 structures. L'analyse conjointe des violations de distance et d'angles dièdres, ainsi que de la liste des pics nOe non attribués en fin de chaque processus a permis d'identifier, puis de corriger, les informations erronées. Ainsi, les pics qui correspondent à du bruit spectral ont été progressivement supprimés, et les erreurs d'attribution des fréquences de résonance de la protéine ont été rectifiées. Le jeu final de données expérimentales RMN retenu a conduit à l'attribution de 3347 pics nOe par le programme d'attribution automatique. Moins de 2% des pics de ce jeu final n'ont pas été attribués. Après application des différents filtres et suppression des nOe redondants, une liste de 2716 contraintes de distance contenant, 2192 contraintes non ambiguës, et 524 contraintes ambiguës, a finalement été établie, ce qui représente une vingtaine de contraintes de distance par résidu (Tableau 4.1). Attribution automatique des pics nOe : Nombre de pics du jeu final 3413 Nombre de pics attribués 3347 Nombre de pics non attribués 66 Nombre de contraintes expérimentales : Contraintes de distance 2743 Non ambiguës 2219 nOe attribués par le programme 2192 nOe de topologie attribués manuellement 9 Distances déduites de l'échange 1H ? 2D 18 Ambiguës (générés par le programme) 524 Contraintes dihédrales et 139 Prédictions TALOS 136 Angles déduits de la constante de couplage 3JHN-Há 3 Tableau 4.1 : Contraintes expérimentales utilisées pour le calcul final de la structure. 1.3.3) Evaluation de la qualité des structures calculées Les 20 structures de plus basse énergie de l'itération finale ont été sélectionnées et soumises à un protocole de raffinement dans le champ de force CHARMM22. A l'issue de cette dernière étape, les 12 meilleurs modèles ont été retenus et constituent l'ensemble final de la structure du domaine K2 résolue par modélisation moléculaire sous contraintes RMN. Le tableau 4.2 présente un résumé des statistiques structurales de ces 12 meilleures structures. Dans l'ensemble, les faibles valeurs des différents termes de la fonction d'énergie suggèrent que les modèles générés sont de bonne qualité. Aucune violation de distance supérieure à 0.5 Å ou d'angle dièdre et supérieure à 10° n'est observée, ce qui indique une bonne prise en compte des contraintes expérimentales RMN. Ces structures présentent également une bonne géométrie covalente avec de faibles valeurs de déviation par rapport à la géométrie idéale des longueurs de liaisons, des angles de valence, et des angles dièdres impropres. Contraintes expérimentales : Contraintes de distance 2743 Contraintes dihédrales et 139 Nombre de violations de distance > 0.5 Å 0 Nombre de violations d'angle et > 10° 0 Energies (kcal/mol) : Liaisons 141 #177; 6 Angles 599 #177; 15 Impropres 20 #177; 2 Van der Waals 146 #177; 15 Electrostatique -671 #177; 25 nOe 143 #177; 10 Dièdres et 5.6 #177; 0.7 Totale 258 #177; 5 Déviations à la géométrie idéale : Liaisons (Å) 0.0159 Angles (°) 3.35 Impropres (°) 2.74 nOe (Å) 0.032 Analyse du diagramme de Ramachandran* Résidus dans les régions favorables 88.6 % Résidus dans les régions additionnelles permises 10.1 % Résidus dans les régions généreusement permises 0.8 % Résidus dans les régions interdites 0.5 % *réalisée avec le logiciel Procheck dans la région 13-107 Tableau 4.2 : Statistiques structurales de l'ensemble des 12 structures finales de K2. La répartition des angles et sur le diagramme de Ramachandran confirme la qualité des structures. Plus de 99 % des valeurs de ces angles occupent les régions énergétiquement favorables ou permises, et seules 0.5 % sont situées dans les régions interdites. La distribution des angles dièdres et pour chaque résidu est présentée en Figure 4.8A. Les éléments de structure secondaire également reportés ont été définis à partir de ces valeurs d'angle. La convergence des modèles générés constitue également un critère de qualité. La superposition des 12 structures finales sur les atomes de la chaîne principale (N, CO, et Cá) montre la bonne définition du squelette peptidique entre les résidus F15 et L106 (Figure 4.7). Pour les acides aminés situés entre ces 2 résidus, la moyenne des écarts quadratiques « rmsd » calculée sur le squelette de l'ensemble final par rapport à une structure moyenne est de 0.43 Å. Cette faible valeur indique une bonne convergence des modèles vers une structure tridimensionnelle unique entre les résidus F15 et L106. N-ter C-ter F15
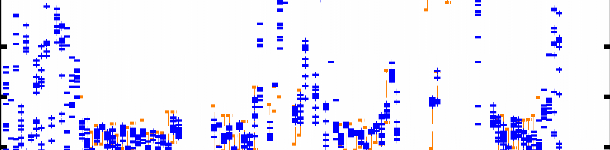
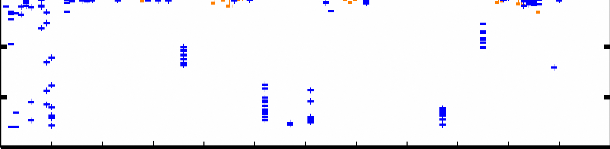
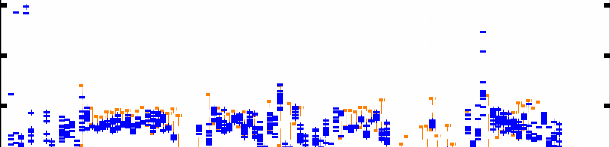
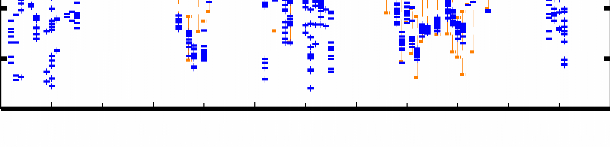
L106 Figure 4.7 : Superposition des 12 structures finales du domaine K2 sur les atomes du squelette (N, CO, et Cá) entre les résidus Q13 et E107. Les hélices á sont représentées en bleu, les feuillets â en rouge, l'hélice 310 en vert pastel, et les régions déstructurées en violet. La Figure 4.8B montre les valeurs des écarts rmsd calculés sur le squelette et les atomes lourds de chaîne latérale pour chaque résidu. On constate une dispersion des angles et associée à de fortes valeurs de rmsd au niveau des extrémités N- et C-terminales G1-P12 et K109-K111. Ceci est en accord avec l'analyse du nOe hétéronucléaire 1H-15N (cf. § 1.2.1) qui indique que ces 2 extrémités sont flexibles et déstructurées. A contrario, la bonne convergence des angles dièdres et les faibles valeurs de rmsd sur les atomes du squelette (< 1 Å) des résidus F15 à L106 suggèrent que ces acides aminés constituent le corps globulaire de la protéine. Ainsi, on observe une bonne définition du squelette peptidique dans les régions structurées en hélice et feuillet, mais également dans les boucles. D'autre part, la majorité des valeurs de rmsd calculées sur les atomes lourds de chaîne latérale sont inférieures à 1.5 Å, ce qui démontre une orientation privilégiée de la plupart des chaînes latérales dans la région F15-L106. La moyenne des écarts rmsd sur les atomes lourds entre les résidus F15 et L106 est de 1.03 Å. Chapitre 4 : Caractérisation structurale du domaine K2 par RMN et Modélisation Moléculaire A) [°] 200 150 100 50 0 -50 -100 -150 -200 H1 S1 H2 310 H3 S2 S3 H4 203 [°] 200 150 100
50 0 -50 -100 -150 -200 10 20 30 40 50 60 70 80 90 100 110 B) 5 4.5 4 3.5 rmsd [Å] 3 2.5 2 1.5 1 0.5 0 10 20 30 40 50 60 70 80 90 100 110 203 0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100 105 110 Figure 4.8 : Analyse de la convergence des 12 structures finales sélectionnées. A) Distribution des angles dièdres et pour chaque acide aminé de ces 12 structures. Les traits oranges correspondent aux intervalles de contrainte d'angle définis dans CNS. B) Déviation quadratique moyenne rmsd pour chaque résidu calculée par rapport à la structure moyenne des 12 modèles sélectionnés (en bleu : superposition sur les atomes N, Cá, et CO du squelette ; en violet : sur tous les atomes lourds de chaîne latérale). Les résidus 36, 74, 77, et 87 sont des glycines. |
|