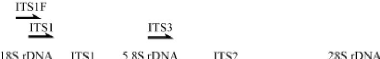Analyse de la diversité des champignons ectomycorhiziens et des ectomycorhizes du raisinier bord de mer (Coccoloba Uvifera l.) le long d'un gradient de salinité en forêt littorale( Télécharger le fichier original )par Raymond AVRIL Université des Antilles et de la Guyane - Master 2 2009 |

2.2. Diversité des ECML'inventaire des ECM est une approche qui permet aussi d'accéder à la composition des communautés fongiques (Agerer, 1991). Basée uniquement sur des caractères morphologiques, anatomiques et histologiques, l'identification des ECM reste très aléatoire. En effet, la couleur du manteau peut changer en fonction de l'âge de la plante hôte ou de l'environnement (Thoen & Bâ, 1989; Wurzburger et al., 2001 ; Diédhiou et al., 2004). Les clés d'identification des ECM proposées jusqu'ici se sont avérées dans l'ensemble insuffisantes pour identifier un champignon à partir d'une ECM à l'exception des ECM typiques du genre Cenococcum (Agerer, 1991). Les caractères macroscopiques et microscopiques des ECM s'avèrent peu concluants pour remonter aux sporophores. De plus, un champignon peut former des ECM dont la morphologie change selon l'hôte ou quelquefois en fonction de l'âge de l'hôte. Des champignons différents peuvent former des ECM morphologiquement similaires. Tout compte fait, l'abondance des sporophores renseigne peu sur la fréquence des ECM. Des champignons comme Suillus fructifient abondamment et forment peu d'ECM. Ë l'inverse, Thelephora forment jusqu'à 40 % des ECM en fructifiant peu ou pas (Diédhiou et al., 2009). Il est rarement possible d'observer des connexions entre le mycélium à la base du pied des sporophores et celui des ECM (Thoen & Bâ, 1989; Rivière et al., 2007). On suppose, souvent à juste raison, que les sporophores, fructifiant au voisinage des arbres, sont potentiellement ectomycorhiziens. Encore faut-il établir un lien formel entre sporophores et ECM car on peut aussi trouver des champignons parasites ou saprophytes à proximité des arbres à ECM. C'est le cas de quelques russules saprophytes facultatives ou parasites (p.ex. Russula parasitica) en Afrique tropicale (Buyck et al., 1996). Pourtant, le genre Russula est souvent ectomycorhizien. L'obstacle principal est l'identification de la composante fongique des ECM, qui peut être surmonté par des méthodes d'écologie moléculaire (p.ex. amplification et séquençage de l'ADNr). 2.3. Caractérisation moléculaire de la diversité fongiqueDifférentes techniques de biologie moléculaire (p.ex. PCR-RFLP, séquençage), basées sur l'analyse de l'ADNr, ont été développées ces dernières années pour étudier la diversité génétique des sporophores, identifier la composante fongique des ectomycorhizes et suivre la persistance des souches fongiques introduites en pépinière et en plantation (Gardes et al., 1991; Martin et al., 1991 ; Selosse et al., 1999; Diédhiou et al., 2004; Rivière et al., 2007; Sanon et al., 2009). La technique PCR est utilisée pour amplifier différentes parties du génome en ayant pour cibles l'ADN total, l'ADNr nucléaire ou l'ADNr mitochondrial. L'ADN total est analysé par des techniques comme l'AFLP (Ç Amplified Fragment Length Polymorphism È), l'ISSR (Ç Inter Simple Sequence Repeat È), la RAPD (Ç Random Amplified Polymorphism DNA È) ou les microsatellites pour accéder au polymorphisme de larges portions d'ADN (Jacobson et al., 1993 ; Redecker et al., 2001 ; Zhou et al., 2001). L'ADNr mitochondrial possède des entités qui sont en plusieurs copies indépendantes du génome nucléaire et utilisées pour des études sur la structuration des communautés de champignons. Le gène de la grande
sous-unité de l'ARNr mitochondrial (<< mtLSU rRNA È) en
particulier le fragment d'environ 450 pb amplifié par les amorces
ML5/ML6, est souvent utilisé en phylogénie des champignons
ectomycorhiziens (Bruns et al., 1998 ; Stendell et
al., 1999 ; Rivière et al., 2007). Bien que cette
région soit peu évolutive au niveau de l'espèce, elle
permet néanmoins de différencier sans ambigu
Figure 2. Représentation schématique de l'ITS (ITS1, ITS2 et 5.8S) de l'unité répétée de l'ADN ribosomique chez les champignons. Les flèches représentent les sites de fixation des amorces les plus utilisées en écologie moléculaire des champignons (Tedersoo et al., 2007). |
|