1- Efficacité d'utilisation de l'eau
Généralement l'efficacité d'utilisation
de l'eau (EUE) exprime le rapport de la matière sèche totale
produite à l'évapotranspiration pendant une période
donnée (Richards & al., 2002) Si cette période est
spécifique au cycle cultural, on parle d'efficacité d'utilisation
nette (EUE nette); si elle s'étale durant toute l'année, on parle
d'efficacité d'utilisation brute (EUE brute) (Bos, 1985). Lorsque la
production considérée n'est pas la biomasse totale, mais une
partie de celle-ci, telles que les grains ou les fruits, on parle alors
d'efficacité d'utilisation de l'eau pour le rendement. D'un point de vue
économique, l'efficacité de l'eau est définie comme le
rapport du rendement économique à la quantité d'eau
utilisée (Gardner& al., 1985).
1-1 Efficacité d'utilisation de l'eau comme
critère de sélection
L'efficacité d'utilisation de l'eau, comme objective de
sélection, peut être définie dans différents
domaines qui dépendent de l'échelle et de l'unité de
mesure considérée (figure 4) sachant que A est le taux de
photosynthèse nette exprimé en umol/m2/s de CO2; T,
est le taux de transpiration exprimé en mmol/m//s d'H2O et
Ä13C est la discrimination isotopique du carbone
exprimée.
|
|
|
|
|
|
|
Echanges gazeux
|
|
|
|
|
Mesurée
|
|
|
|
|
|
|
|
|
|
Intégrée
|
|
|
|
|
|
|
|
|
EUE
|
|
EUE instantanée = A/T
EUE intrinsèque = A/g
EUE biomasse =
matière
sèche/évapotranspiration
EUE rendement= rendement
grains/évapotranspiration
|
Estimée
|
|
|
|
Discrimination Isotopique du
Carbone
|
|
|
|
|
|
|
Figure 4. Echelle d'étude de l'efficacité
d'utilisation de l'eau (Tambussi, & al., 2007).
Toutes les définitions potentielles ont en commun: un
échange d'eau contre une unité de production.
Pour les fermiers et les agronomes, l'efficacité
d'utilisation de l'eau correspond au rapport de la matière sèche
produite à l'évapotranspiration de la culture (EUE),
exprimée par la relation suivante:
EUE =W/[1 +(Es/T)] (1)
Où W est l'efficacité de la transpiration
(matière sèche/transpiration), Es est l'eau perdue par
évaporation de la surface du sol et T est l'eau perdue par transpiration
du végétal (Richards, 1991).
La relation (1) montre que l'efficacité d'utilisation
de l'eau peut être améliorée soit par, l'augmentation de
l'efficacité de la transpiration, soit par une réduction de
l'évaporation du sol. L'importance relative de chacune des composantes
de EUE varie selon la distribution des pluies. Si les pluies sont rares pendant
la saison de croissance, et que la culture utilise uniquement l'eau
stockée dans le sol, alors l'augmentation de l'efficacité de la
transpiration (W) offre une belle opportunité pour l'amélioration
de EUE. En revanche, si l'alimentation en eau de la culture repose uniquement
sur les pluies, alors la réduction de l'évaporation du sol (Es)
fournit un autre moyen pour l'amélioration de EUE (Richards &
al., 2002).
Pour les physiologistes, l'unité de base de la
production est le gain d'un mole de carbone dans la photosynthèse (A)
par rapport à l'eau transpirée (T). Donc, cette définition
correspond à l'EUE instantanée à l'échelle des
échanges gazeux de la feuille (A/T). Ces deux définitions peuvent
être reliées par l'équation suivante (Passioura, 1977):
Rendement = ET * T/ET * W * HI (2)
Dans cette relation, le rendement est défini comme
étant une fonction du produit de; l'eau utilisée par la culture
(évapotranspiration ET), la proportion de la transpiration actuelle de
la culture (T/ET), l'efficacité de la transpiration pour la production
de biomasse (W) et qui correspond à la quantité de biomasse
produite par millimètre d'eau transpirée et enfin le ratio entre
le rendement en grain et la biomasse aérienne (HI) (Condon &
al., 2004).
Dans la relation (2), aucune des composantes n'est vraiment
indépendante l'une des autres (Condon & Richards, 1993), mais
chacune d'elles peut faire l'objet d'amélioration
génétique. L'efficacité d'utilisation de l'eau à
l'échelle foliaire, A/T, est directement et uniquement liée
à l'une de ces composantes, W, qui est l'efficacité de la
transpiration pour la production de biomasse. Potentiellement donc, A/T peut
influencé l'une des trois autres composantes (Condon & al.,
2004).
1-2 Sélection pour l'efficacité
d'utilisation de l'eau à l'échelle de la feuille
La recherche pour l'amélioration de l'efficacité
d'utilisation de l'eau agronomique par la sélection pour une grande
efficacité d'utilisation de l'eau à l'échelle de la
feuille a été longtemps attractive (Fischer, 1981).
A/T peut être décrite mathématiquement en
notant en premier que A est le produit de la conductance stomatique pour le
CO2, Gc et le gradient de la concentration du CO2 entre l'extérieur (Ca)
et l'intérieur (Ci) de la feuille (équation 3):
A = Gc(Ca- Ci) (3)
Deuxièmement, T est le produit de la conductance
stomatique de la vapeur d'eau Gw et le gradient de la concentration de la
vapeur d'eau, à l'intérieur (Wi) et l'extérieur (Wa) de la
feuille (équation 4)
T = Gw(Wi-Wa) (4)
Puisque la concentration du CO2 est plus grande à
l'extérieur de la feuille, alors que celle de l'eau est plus grande
à l'intérieur, le ratio A/T devient donc équation (5):
A/T = [Gc (Ca- Ci)] /
[Gw(Wi-Wa)] (5)
L'équation 4 peut être simplifiée par
l'équation 6, sachant que le ratio entre la diffusion du CO2 et de l'eau
dans l'air est égale à 0,6:
A/T 0,6 Ca(1 - Ci/Ca) / (Wi-Wa)
(6)
L'équation (6) indique deux voies possible pour
l'amélioration de l'efficacité d'utilisation de l'eau à
l'échelle de la feuille : soit, par l'abaissement du rapport Ci/Ca et
donc l'augmentation de la valeur de (1-Ci/Ca). Soit, de rendre la valeur de
(Wi-Wa) plus petite, par diminution des pertes en eau en abaissant le gradient
de la transpiration
1-2-1 Diminution du gradient de la concentration en
vapeur d'eau
Le moyen le plus simple par lequel la sélection a
amélioré l'efficacité de la transpiration pour la
production de biomasse via A/T, a été la modification des
caractéristiques des cultures, comme la réduction du gradient
d'évaporation durant le cycle de croissance de la culture (Tanner &
Sinclair, 1983; Richards & al., 2002).
Ainsi, le processus de réflectance au niveau des
feuilles diminue la perte en eau, de la culture provenant du gradient de la
concentration de la vapeur d'eau entre le couvert végétal et
l'atmosphère. Ce gradient est faible dans les régions froides,
humides et dans la plupart des régions durant les mois les plus froids
de l'année.
Durant le dernier siècle, les sélectionneurs de
nombreuses espèces cultivées, ont exploité la variation
génétique associée à la précocité, la
réponse à la photopériode et au besoin en vernalisation,
pour générer d'énormes variations dans la
phénologie des cultures. Cette variation phénologique a permis
aux cultures de croître successivement dans des régions et
à des moments de l'année ou la demande évaporative
prévalente est faible, ce qui engendre une élévation de
A/T et une augmentation des rendements des cultures.
Pour saisir cette opportunité, tous les efforts doivent
être axés pour une amélioration de l'efficacité
d'utilisation de l'eau des cultures. L'ajustement de la date de semis pourrait
améliorer le ratio A/T et donc le rendement des cultures (Condon &
al., 2004).
1-2-2 Changement du rapport Ci/Ca
En référence à l'équation (6), une
autre voie est offerte à la sélection pour l'amélioration
de A/T et de ce fait à l'amélioration de l'efficacité de
la transpiration pour la production de biomasse. Il s'agit d'élever le
numérateur du rapport (1-Ci/Ca) pour sélectionner des
génotypes qui ont une faible valeur de Ci/Ca.
La faible valeur de Ci/Ca peut refléter une valeur
faible de la conductance stomatique, une grande capacité
photosynthétique ou la combinaison des deux (Farquhar & al.,
1989).
Théoriquement, la réduction de Ci/Ca de 0,7
à 0,6 aboutit à un gain de 33% d'efficacité d'utilisation
de l'eau, sachant que cette dernière est proportionnelle à la
valeur (1-Ci/Ca). Cependant, des disfonctionnements peuvent avoir
lieu. Par exemple, si la baisse de la valeur de Ci/Ca est due
à une augmentation de la capacité photosynthétique, ceci
conduit à une augmentation de l'assimilation du CO2 pour chaque
unité de mesure, et si la baisse de (Ci/Ca) est due à une faible
conductance stomatique, alors elle conduit à une diminution de
l'assimilation du CO2.
Outre la diminution de l'assimilation du CO2, une autre
pénalité accompagne la conductance stomatique : si la conductance
de la couche externe de la feuille n'est pas très élevée,
la température foliaire et la concentration en vapeur d'eau à
l'intérieur de la feuille (Wi) augmentent et en parallèle la
concentration en vapeur d'eau à l'extérieur de la feuille et la
conductance baissent. Les modifications de Wi et Wa aboutissent
à l'augmentation de la transpiration par unité de conductance
stomatique, et par conséquence à l'augmentation de
l'efficacité d'utilisation de l'eau comme prédite par
l'équation (6) (Condon & al., 2002,2004).
2- Discrimination isotopique du carbone 2-1 Relation
entre ?13C et le ratio Ci/Ca
Il est important de noter que la conductance stomatique, la
capacité photosynthétique et A/T sont très difficiles
à mesurer malgré la disponibilité des systèmes de
mesure des échanges gazeux au niveau de la feuille. Actuellement, il est
admis qu'au moins chez les espèces à photosynthèse de type
C3, des différences relatives de Ci/Ca, peuvent être
estimées indirectement, en mesurant la composition isotopique du carbone
de la matière sèche de la plante (Farquhar & al.,
1989).
Le 13C est un isotope stable, il constitue
près de 1% du CO2 atmosphérique. Chez les plantes C3, la
proportion du 13C est plus petite que celle de l'atmosphère,
car, les espèces de type C3 discriminent contre le 13C
pendant la photosynthèse. La discrimination isotopique du carbone
(?13C) est une mesure du ratio 13C/12C dans le
matériel végétal, relativement au même ratio de
l'air qui l'entoure et d'où elle puise le CO2 .et qui peut être
défini comme suit (Farquhar & Richards, 1984):
?13C = [(Ra/Rp) - 1] * 1000 (7)
ou Ra est la valeur du ratio
13C/12C dans l'atmosphère et R p est
celle du matériel végétal. Pour des raisons de
commodité l'unité de ?13C est exprimée en (%o).
Chez les plantes de type C3, le ratio C13/C12 a une
valeur proche de 1,02 donnant une valeur de ?13C proche de 20%o.
Chez les espèce C3, de nombreux processus contribuent
à la détermination de la valeur de ?13C mesurée
dans leur matière sèche (Farquhar & al., 1989;
Brugnoli & Farquhar, 2000).
L'expression approximative suivante (équation 8) a
été développée par Farquhar et Richards (1984) pour
expliquer deux processus majeurs; le fractionnement associé à la
diffusion du CO2 dans les espaces intercellulaires (a = 4,4%o) et le
fractionnement associé à la carboxylation du CO2 par la Rubisco
dans les produits primaires de la photosynthèse (b 28%o)
Ä13C = a + (b - a)Ci/Ca (8)
La valeur 28%o de b est une approximation parce qu'il existe
des sources supplémentaires de la variation de Ä13C,
telles que celles associées au métabolisme et à la
respiration et qui restent mal définies et dont la magnitude est faible
dans la plus part des situations (Brugnoli & Farquhar, 2000). Les valeurs
numériques citées dans la formule (8) donnent:
Ä13C 4,4 + 23,6 Ci/Ca (9)
Cette simple équation montre que Ä13C
est positivement corrélée à Ci/Ca. Comme
auparavant il a été admis que A/T devrait être
corrélée négativement avec Ci/Ca (équation 6),
alors, Ä13C et A/T doivent être toujours
négativement corrélées.
2-2 Relation entre ?13C et l'efficacité
d'utilisation de l'eau
Le fait d'admettre que Ä13C pourrait fournir
une mesure relativement simple et indirecte de la variation du rapport A/T
(Farquhar & al., 1982; Farquhar & Richards, 1984) a
relancé la perspective de l'exploitation de la variation de
l'efficacité d'utilisation de l'eau à l'échelle foliaire
pour améliorer l'efficacité d'utilisation de l'eau.
Cependant, Il a été montré dans d'autres
travaux, qu'il existe une légère variation de Ci/Ca parmi les
espèces de type C3, et qu'elle est l'unique différence
substantielle entre les espèces de type C3 et C4 (Fischer, 1981; Tanner
& Sinclair, 1983). Depuis des études novatrices ont mis en
évidence pour plusieurs espèces C3 que la variation en
Ä13C reflète étroitement la variation dans le
ratio Ci/ Ca (Farquhar & al., 1982; Farquhar&
Richards, 1984).
Brugnolli et Farquhar (2000), ont mesuré
?13C in situ à partir du courrant d'air à
l'entrée et à la sortie de la feuille ainsi qu'au niveau des
photosynthétats extraits fraîchement des feuilles ou mesuré
dans la matière sèche des tissus. La variation de ?13C
entre les génotypes des espèces de type C3 est assez importante.
Théoriquement, cette variation est substantielle pour A/T et pour
l'efficacité d'utilisation de l'eau ainsi que pour le potentiel de
production de la matière sèche (Farquhar et Richards, 1984).
Des corrélations négatives entre ?13C
et l'efficacité d'utilisation de l'eau par les plantes ont
été enregistrées chez plusieurs espèces (Farquhar
& al., 1989; Hall & al., 1994; Condon & Hall,
1997). Depuis, ?13C est considéré comme un
caractère à forte héritabilité qui est relativement
facile à manipuler dans la sélection des populations (Condon
& Richards, 1992; Rebetzke et al., 2002).
La mesure de ?13C peut être effectuée
sur des échantillons de matériel végétal frais ou
stocké, c'est est une technique relativement rapide qui peut être
automatisée. Cependant, de sérieuses difficultés sont
survenues lorsqu'on a essayé de passer de l'association entre
Ä13C et l'efficacité d'utilisation de l'eau des feuilles
et des plantes entières vers l'association entre Ä13C et
l'efficacité d'utilisation de l'eau et le rendement obtenu au champ. La
nature de ces difficultés et les solutions possibles sont
abordées dans les points suivants.
2-3 Relation entre rendement en grains et ?13C
chez les céréales
La difficulté majeure à laquelle se heurte
l'utilisation de la discrimination isotopique du carbone ?13C dans
la sélection pour l'amélioration agronomique de
l'efficacité d'utilisation de l'eau est l'incohérence
observée entre ?13C et le rendement. Ces incohérences
ont été bien répertoriées dans de nombreuses
études impliquant les céréales, tels que le blé
tendre (Triticum aestivum L.), le blé dur (T. turgidum
var. D.) et l'orge (Hordeum vulgare L.).
L'association négative entre ?13C et A/T
à l'échelle foliaire et les associations négatives
observées entre ?13C et l'efficacité d'utilisation de
l'eau à l'échelle de la plante dans plusieurs études
menées dans des pots, ont toujours révélé des
corrélations négatives entre le rendement des cultures et
?13C.
Aussi, beaucoup d'études sur des collections de
céréales cultivées, en Australie sous régime
pluvial ou irrigué et dans d'autre régions (Sayre & al.,
1995; Fischer & al., 1998), ont montré que les
relations entre le rendement en grains et ?13C ont été rarement
négatives, si ce n'est souvent des relations positives ou neutres.
Beaucoup de ces études associant ?13C et la productivité chez les
céréales ont utilisé des groupes de génotypes qui
diffèrent entre eux non seulement par ?13C, mais aussi par la date de
floraison et la hauteur, deux caractéristiques qui peuvent
influencées considérablement les productions,
indépendamment de ?1 3C (Condon & al., 1987, 1993, 2002;
Condon & Richards, 1993; Condon & Hall, 1997), en région
méditerranéenne (Voltas & al., 1999; Merah &
al., 2001; Royo & al., 2002; Araus & al.,
2003)
Il existe plusieurs raisons qui peuvent expliquer que de
nombreuses études ont montré des relations variables entre le
rendement et ?13C. Chez les céréales un faible
?13C (donc un fort A/T) est un caractère dominant en
matière l'utilisation de l'eau, probablement plus important en
matière de taux de croissance du végétal. En effet, en
l'absence de déficit hydrique du sol, les génotypes à
faible ? 13C ont tendance à croître moins vite que ceux
à fort ?13C, et par conséquent, ils produisent moins
de biomasse et de rendement grain (Condon & Richards, 1993; Condon &
al., 1993, 2002; Lopez-Casteneda & al., 1995).
Une raison évidente largement citée fait que
chez les cultures de céréales, ?13C est associé
au caractère dominant du taux de croissance. En effet, la
différence dans ?13C en absence de déficit hydrique du
sol résulte de la variation de la conductance stomatique. Les
génotypes à faible conductance stomatique ont tendance à
avoir un fort A/T et un faible ?13C. Dans ce cas, A/T sera
vraisemblablement associé à un faible taux de
photosynthèse par unité de surface foliaire et un taux de
croissance faible.
La faible conductance, n'est pas la seule raison qui explique
l'association entre A13C et le caractère dominant du taux de
croissance de la culture. Chez les céréales, la variation de
A13C peut résulter aussi de la variation da la
capacité photosynthétique (Condon & al., 1990;
Morgan & Le Cain, 1991). Si la faible valeur de A13C est le
résultat d'une forte capacité photosynthétique, alors il y
aura un fort taux de photosynthèse par unité de surface foliaire
et donc un taux de croissance de la culture plus rapide. Les augmentations
substantielles de la capacité photosynthétique sont facilement
réalisées grâce à une concentration en azote dans
les petites feuilles qui interceptent moins la lumière. Cette faible
interception peut ralentir le taux de croissance (Condon & Richards, 1993;
Condon & Hall, 1997). Si l'interception de la lumière n'est pas
réalisée ou l'est promptement, comme dans les cultures en sec,
alors la forte capacité photosynthétique ne va pas entrainer une
augmentation de la croissance.
Par ailleurs, des génotypes de céréales
à faible capacité photosynthétique peuvent avoir une
croissance plus rapide, une corrélation positive entre la production de
biomasse à l'anthèse et A13C .Dans une
expérience où ont été testés trois
génotypes ayant de fortes valeurs de A13C et de faibles
valeurs de conductance stomatique, Il a été montré que
pour des conductances similaires mais avec de fortes valeurs de
A13C, les génotypes avaient une faible capacité
photosynthétique. Cette déduction a été
confirmée sur de grandes plantes cultivée en serre (Condon &
al., 1990).
Ainsi, chez les céréales, une grande
capacité photosynthétique ne peut être
nécessairement associée à un plus fort taux de croissance
de la culture. Cette conclusion peut être confortée par
l'étude de Fischer & al., (1998) sur les changements
physiologiques du blé tendre publié par le CIMMYT de 1962
à 1988. Ces blés récents ont relativement un grand
rendement, mais, il n'est pas lié à une production plus grande de
la biomasse. Par ailleurs, ce gain en rendement reflètent un plus fort
indice de récolte pour la plus part de ces blés récents.
Parmi l'historique collection des blés du CIMMYT, il n' y a pas eu de
changement de production de la biomasse total malgré que les blés
récents aient une plus grande capacité photosynthétique et
une forte conductance stomatique, qui à elles seules
génèrent relativement une grande valeur de A à
l'échelle de la surface foliaire.
3- Interaction entre croissance et utilisation de
l'eau
Indépendamment de sa base physiologique, chez les
génotypes des céréales, le caractère dominant de la
croissance à travers un faible A13C, a des implications
importantes pour l'efficacité agronomique de l'utilisation de l'eau. Les
plus importantes relations entre A13C et le rendement ont
été observées dans des milieux ou des saisons durant
lesquels, les irrigations complémentaires ou les précipitations
régulières, maintenaient un statut hydrique élevé
du sol durant le cycle de croissance. Dans ces environnements, la rapide
croissance des génotypes à forte discrimination isotopique du
carbone, se traduit souvent et directement par, une plus grande production de
biomasse finale et de rendement en grains (Condon & al., 1987,
2002; Sayre & al., 1995; Fischer & al., 1998).
A maturité, les génotypes à faible
A13C qui produisent moins de biomasses et de rendements,
n'exploitent pas l'eau restante dans le sol (Condon & al.,
2002).
Dans les environnements moins favorables, une limitation en
eau suite à une variation en quantité où dans le temps,
peut interagir avec le caractère dominant de la croissance et de
l'utilisation de l'eau des génotypes à faible A13C,
pour générer des relations complexes entre le rendement et
A13C. Cette complexité peut être illustrée par
l'étude de Condon & Hall, (1997) sur deux lignées F6 et F7
cultivées durant 1992 et 1993. Pour l'année sèche de 1992,
ils ont observé, chez le premier croisement (Rosella x Matong), une
liaison entre A13C et la faible production de biomasse et de
rendement, mais chez le deuxième (Quarrion x Cranbrook) il n'y avais pas
de liaison entre la productivité et A13C. Le statut hydrique
du sol était vraisemblablement élevé pour le second
croisement, parce que la deuxième lignée a fleurit une semaine en
avance par rapport à la première.
Les résultats de ces deux lignées
cultivées sur ces deux saisons, indiquent que la quantité des
pluies est une variable importante qui contribue dans la variation des
rendements en grains. Aussi, ils indiquent que le développement d'un
déficit hydrique du sol correspondant à la phase critique de la
floraison est aussi une variable importante.
Si les génotypes à fort A13C
épuisent l'eau disponible dans le sol très rapidement avant la
floraison, ce sera une pénalité pour leur rendement (Fischer,
1981). Cependant, la pénalité en matière de rendement et
d'utilisation de l'eau par la culture, associés à la rapide
croissance des génotypes de céréales à forte
A13C, n'est pas aussi grande à celle prévue. En fait,
la croissance rapide des génotypes de céréales à
fort A13C, a toujours été considérée
comme avantageuse dans les environnements ou pendant les saisons où les
fréquences de chute de pluies précoces dans la saison sont
très importantes, mais ceci n'est pas le cas pour les stades
avancées en climat méditerranéen typique (Condon &
al., 1993; Condon & Hall, 1997; Voltas & al., 1999;
Merah & al., 2001; Royo & al., 2002; Araus &
al., 2003).
Dans ce genre d'environnement, l'évaporation de la
surface du sol représente près de 50% des précipitations
de la saison de culture (Cooper et al., 1987). Les études,
où l'eau utilisée a été bien répartie entres
les plantes et le sol, ont montré que la plus forte transpiration
associée au fort A13C résulte de la différence
d'utilisation de l'eau total jusqu'à l'anthèses bien que les
génotypes à fort A13C aient une croissance plus rapide
à ce critique stade de développement (Condon & al.,
1993, 2002; Lopez-Castaneda & al., 1995). La raison de ceci
est que, contrairement aux génotypes tardifs à faible
A13C, les génotypes à fort A13C et à
croissance précoce, arrivent à couvrir rapidement le sol, et par
là, à bien réduire l'évaporation du sol lorsque les
chutes de pluies sont fréquentes. Parce que l'eau total utilisée
jusqu'à l'anthèse était similaire, les génotypes
à fort A13C produisent plus de biomasse et arrivent à
la transloquer pour produire un nombre élevé de grains et de
rendement.
Les génotypes à fort ?1 3C peuvent produire plus
de biomasse à l'anthèse en puisant la réserve en eau du
sol (Condon and al., 1993). La faible croissance après la
floraison comparée aux génotypes à faible ?13C qui sont
plus économes dans l'utilisation de l'eau. Les génotypes à
fort ?1 3C donnent plus de rendement, grâce à la production d'un
grand nombre de grains (associée à une plus grande production de
biomasse à l'anthèse) et grâce à leur
capacité à les remplir, probablement par translocation de grandes
quantités d'assimilats stockées. Dans les mêmes
environnements, les génotypes à faible ?13C n'arrivent pas
à utiliser efficacement les assimilats stockées. Ceci est
dû à leurs faibles réserves engendrées par une
moindre production de biomasse à l'anthèse, ou bien les
assimilats stockées n'étaient pas utiles, car les
génotypes à faible ?13C avaient plus de réserves hydriques
dans leurs sol dont ils se servaient pour le remplissage des grains grâce
à une forte photosynthèse (Condon and Hall, 1997). En
dépit de leurs faibles croissances après la floraison, les
génotypes à fort ?13C réalisent des rendements
élevés, car ils ont tendance à avoir un indice de
récolte très élevé.
Pour résumer la complexe association entre
?13C, la croissance, l'utilisation de l'eau et le rendement chez les
céréales, le caractère conservatif de la croissance
associé avec le faible ?13C et la forte A/T a
été développé pour interagir avec l'ampleur et le
moment des limitations en eau, afin de générer des effets divers
sur la structure des composantes du rendements précédemment
citée (équation 1).
Dans certaine études, une grande efficacité de
transpiration pour la production de biomasse (W), reflétant une forte
A/T à l'échelle foliaire, a été compensée
par une utilisation réduite de l'eau totale de la culture. Les
génotypes à fort ?13C ont donné plus de
rendement et ont épuisé toute l'eau contenue dans le profil du
sol. Les génotypes à faible ?13C extraient moins d'eau
et perdent plus d'eau par évaporation du sol. Dans la combinaison de ces
deux facteurs (moins d'extraction et plus d'évaporation), quelques fois
il a été observé chez les génotypes à faible
?13C, une transpiration considérablement faible
comparée à celle des génotypes a fort ?13C
(Condon & al., 1993, 2002).
Donc souvent, il n'y a pas eu de relation entre le gain en
biomasse et une forte A/T, et fréquemment entre la production de
biomasse et le faible rendement. La forte production de biomasse chez les
génotypes à fort ?13C a été le plus
souvent réalisée à l'anthèse, cette
différence à l'anthèse a été
substantiellement reflétée par un nombre de grains et un
rendement élevés chez les génotypes à fort
?13C bien qu'ils aient produit moins de biomasse à
après l'anthèse. Tout en prenant en compte le rendement, dans
quelques études, les relations positives entre la production de biomasse
et ?13C ont été soutenues par les relations positives
entre l'indice de récolte (HI) et ?13C qui est probablement
due, en partie, à une forte translocation des réserves
stockées chez les génotypes à fort ?13C
1- Protocole expérimental
L'expérimentation a été
réalisée durant la campagne agricole 2006/2007. L'essai a
été mené sur le site de la pépinière
Latreche Amar, située au lieu dit: `Chirhoum', à 12
kilomètres au Nord Est du chef lieu de la wilaya de Sétif. La
parcelle expérimentale se trouve à une altitude de 1180
mètres, avec une latitude de 36° 10 et 5° 20 de longitude, ses
coordonnées géographiques ont été mesurées
à l'aide d'un GPS `Garmin 72'.
Le précédent cultural est une jachère
labourée au printemps avec une charrue à disque et
recroisée au cover croop en automne, suivi d'un amendement
phosphaté avec du TSP à 46% à raison de 1qx/ha. Le lit de
semence a été préparé par le passage d'une herse
juste avant le semis. On a pratiqué un désherbage à la
main tout au long du cycle de la culture, complété par un
désherbage chimique au stade fin tallage et début montaison avec
du Granstar (12 g/h) et Hussar (1 Litre/h). Une fertilisation azotée a
été réalisée à ce même stade avec de
l'urée à 46% à raison de 1qx/h. Le semis a
été exécuté à la main le 26 novembre 2006
à raison de 75 grains/ml et la récolte a été
réalisée les 28 et 29 juin 2007.
Le dispositif expérimental adopté est en bloc
complètement randomisé à quatre répétitions.
La parcelle élémentaire comporte quatre lignes de 2,50
mètres avec un écartement entre lignes de 18 cm. Pour
éviter l'effet de bordure lors des notations, on a fixé
dès le semis quatre stations de 01 ml par parcelle
élémentaire sur les deux lignes du milieu en écartant les
10 et 30 centimètres, respectivement des extrémités et du
milieu. Le matériel végétal est composé d'un groupe
de 08 variétés dont les caractéristiques sont
données en annexe 1.
Notant enfin que le premier bloc a été
réservé pour le prélèvement d'échantillon
servant pour l'estimation de la matière sèche et du tallage
herbacé, les trois autres blocs ont servi pour l'estimation des autres
paramètres.
2- Notations 2-1 Rendement et composantes du
rendement
Les notations ont été effectuées par
repérage, des dates et le nombre de jours, qui correspondent aux quatre
stades repères du cycle de croissance du matériel
végétal étudié. Il s'agit des stades :
levée, montaison, épiaison et maturité.
Nous avons comptabilisé la phase située entre
50% de la levée et 50% de l'épiaison, puis transformé le
nombre de jours calendaires correspondant, en somme de température par
cumul des températures moyennes journalières positives.
Au stade deux noeuds, nous avons prélevé un
échantillon de 10 plants/génotype qu'on a pesé,
passé à l'étuve à 120 °C puis repeser pour
calculer la matière sèche. Aussi, on a prélevé un
autre échantillon de 10 plants/variété qui a servi pour
calculer le coefficient de tallage herbacé.
Pour les autres blocs, sur toutes les stations nous avons
noté le nombre de plant levé. A maturité, nous avons
récolté les stations séparément. Sur chacune
d'elle nous avons estimé la biomasse aérienne,
la hauteur, le nombre d'épis. La fertilité a été
calculée sur un échantillon de 05 épis de chaque station.
Ces mêmes grains ont été pesés et à l'aide de
la règle de trois nous avons calculé le poids de mille grains. Le
rendement final a été calculé par peser des grains sur
chaque station.
2-2 Teneur en chlorophylle
Sur le milieu de la feuille drapeau de dix plants de chaque
variété, nous avons mesuré la teneur moyenne en
chlorophylle total (a+b), en utilisant un chlorophyllo mètre digital de
type SPAD 501 qui donne des lectures en unité SPAD. Ensuite nous avons
prélevé deux feuilles, dont les teneurs en chlorophylle en
unité SPAD sont connues, et grâce à la technique du DMSO
(Arnon, 1949), nous avons estimer leur teneur en chlorophylle en umol/gramme
puis transformé les unités SPAD en unité umol/g de
matière sèche. On note que pour chaque variété on a
effectué quatre mesures étalées entre les stades
épiaison et remplissage
2-3 Discrimination isotopique du carbone
(Ä)
Pour chaque parcelle élémentaire, on a
broyé finement quelques graines puis pris un échantillon de 02
grammes qui a servi pour mesurer le rapport isotopique, R =
C13/C12, par spectrométrie de masse
(Spectrométrie de masse Optima, GV Instruments, Manchester, UK) dans les
laboratoires de Seibersdorf à l'Agence Internationale à l'Energie
Atomique de Vienne, Autriche.
La valeur de la composition isotopique ä13C(%o)
est obtenue par la relation suivante: ä13C(%o) = [(R
échantillon/R standard)-1]*100 (1)
Le R standard est celui de Pee Dee Belemnite "PDB". Enfin pour
obtenir les valeurs de la discrimination isotopique (Ä), on a
utilisé la relation suivante:
Ä (%o) = [ (ä13Ca -
ä13Cp)/(1 + ä13Cp) ] * 1000 (2)
a et p sont respectivement l'air et l'échantillon
considéré
2-4 Etat hydrique du sol
Afin de caractériser l'état hydrique de notre
parcelle, nous avons réalisé un profil pédologique sur une
profondeur d'un mètre. Il ressort que notre sol présente les
caractéristiques suivantes :
* Une bonne structuration en surface ainsi qu'en subsurface,
collant et peu plastique.
* la structure est de type grumeleuse à polyédrique
respectivement vers la surface et en profondeur.
* sol apparemment jamais inondé mais qui renferme des
matériaux indurés (graviers et petits cailloux) bien
ancrés dans l'ensemble du profil.
La succession lithologique et les processus pédologiques,
laissent penser qu'une accumulation d'origine alluvionnaire s'y est
installée.
Les résultats de L'analyse granulométrique
réalisée (laboratoire de pédologie de l'UFA Sétif)
avec la méthode de la pipette robinson, sur les quatre premiers horizons
avec un pas d'échantillonnage de 20 centimètres, montrent, selon
le diagramme de l'USDA, que la texture est de type Limono argileuse avec une
teneur en sable légèrement élevé pour le premier
horizon (tableau 7). La densité apparente, réalisée avec
la méthode du cylindre, révèle que les horizons
différent entre eux et que le deuxième horizon est plus dense
comparer aux autres.
Tableau 7. Granulométrie et paramètres
hydriques du sol de l'expérimentation.
|
Horizon
(cm)
|
Argile
(%)
|
Limon
Grossier
(%)
|
Limon
Fin
(%)
|
Sable
(%)
|
Densité
Apparente
(g/cm3)
|
Cc
(mm)
|
pF
(mm)
|
RU
(mm)
|
RFU
(mm)
|
|
0-20
|
32,00
|
28,80
|
22,00
|
17,20
|
1,32
|
71,28
|
34,32
|
36,96
|
24,64
|
|
20-40
|
34,90
|
25,05
|
24,70
|
15,35
|
1,42
|
76,68
|
36,92
|
39,76
|
26.51
|
|
40-60
|
34,90
|
25,05
|
24,70
|
15,35
|
1,36
|
73,44
|
35,36
|
38,08
|
25,39
|
|
60-80
|
34,90
|
25,05
|
24,70
|
15,35
|
1,36
|
73,44
|
35,36
|
38,08
|
25,39
|
|
0-80
|
31,38
|
25,99
|
24,03
|
15,8
|
1,38
|
294,8
|
141,9
|
152,8
|
101,9
|
On a utilisé le tableau de correspondance entre texture
et réserve en eau par unité volumique mis en point par l'INRA
(Jamagne & al., 1977), pour estimer la réserve en eau du
sol. Il ressort que la capacité au champ de notre sol est de 27% et son
point de flétrissement est de 13%.
On a pratiqué la méthode gravimétrique
(utilisation d'une tarière agronomique de 120 centimètres) pour
le suivi du profil hydrique du début jusqu'à la fin du cycle
cultural. Pour chaque horizon, Nous prélevons un échantillon de
100 grammes de sol frais, après l'avoir passer à l'étuve
à 105 C° pendant 24 heures, nous le repesons et calculons son
humidité pondérale selon la formule suivante:
Humidité pondérale (%) = (Poids frais - Poids sec)
/ Poids sec 3- Traitement statistique
Concernant l'analyse de variance les données ont
été traitées par le logiciel ViSta (Young, 2001). La
comparaison des moyennes des paramètres mesurés ainsi que les
corrélations et les régressions ont été
réalisées à l'aide du logiciel STATISTICA (StatSoft,
1997).
1- Résultats
1-1 Bilan hydrique par la méthode
gravimétrique 1-1-1 Températures
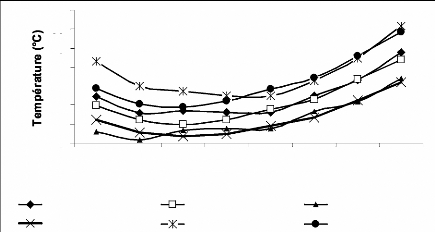
L'examen des données (figure 5) montre que la campagne
2006/2007 a été plus chaude comparée aux dernières
25 années. Seules les températures moyennes mensuelles de Mars
(8,10°C) et Mai (16,56 °C) semblent être proches de la moyenne
des mêmes mois de 1981-2005 avec respectivement 8,94 et 16,71 °C
(tableau 8).
Pour les mois de Novembre et Décembre, on note une
diminution des températures minimales de 3,27 et 2,05 °C
respectivement, suivie d'une élévation des températures
maximales de 7,10 et 4,66 °C. Le mois de Janvier qui est normalement le
plus froid, a enregistré durant cette compagne, une
élévation de température moyen de 3,42 °C, cela est
due à l'augmentation des températures maximales soit ; 4,15
°C en plus par rapport à la normale.
Notons enfin que la vague de chaleur qui a
caractérisée la période allant du 17 au 25/06/2007 (annexe
2a), avec une moyenne de 29,83 °C et un maxima de 37,16 °C,
enregistre une élévation de température de 1.93 °C
pour le moins de Juin comparée aux 25 dernières années.
Tableau 8. Températures (en °C) et
précipitations (en mm) des périodes
1981/2005 et
2006/2007
|
Mois
|
T moy
2006/2007
|
T moy
1981/2005
|
T max
2006/2007
|
T max
1981/2005
|
T min
2006/2007
|
T min
1981/2005
|
Pluie
2006/2007
|
|
Nov
|
12,27
|
9,81
|
2,93
|
6,2
|
21,60
|
14,5
|
6,90
|
|
Déc
|
7,86
|
6,07
|
0,75
|
2,8
|
14,96
|
10,3
|
61,80
|
|
Janv
|
8,45
|
5,03
|
3,36
|
1,7
|
13,55
|
9,4
|
9,50
|
|
Févr
|
8,10
|
6,18
|
3,69
|
2,3
|
12,51
|
11,1
|
23,20
|
|
Mars
|
8,10
|
8,94
|
3,74
|
4,6
|
12,46
|
14,3
|
108,60
|
|
Avr
|
12,38
|
11,56
|
8,34
|
6,8
|
16,42
|
17,2
|
95,60
|
|
Mai
|
16,56
|
16,71
|
10,83
|
11,3
|
22,28
|
23
|
27,80
|
|
Juin
|
23,93
|
22,02
|
17,08
|
16,1
|
30,78
|
29,3
|
22,70
|
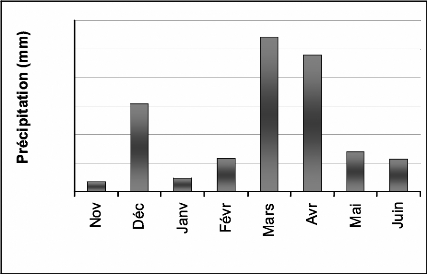
1-1-2 Précipitations
Durant la campagne 2006/2007, 50 jours de
précipitations ont été enregistrés (annexe 2b) dont
deux jours de neige qui ont coïncidé avec le début du
printemps (21 et 22 Mars). Le cumul des précipitations de Novembre
à Juin 2007 a atteint 356,1 mm. Novembre, Janvier et Juin sont les moins
pluvieux, respectivement : 6,9; 9,50 et 22,70 mm (tableau 8). Décembre a
enregistré 61,80 mm, alors que le pic a été
enregistré au mois de Mars et Avril, qui totalisent plus de la
moitié des précipitations cumulées durant le cycle
cultural (figure 6).
Nov Déc Janv Févr Mars Avr Mai Juin
Mois
Tmoy2006/2007 Tmoy 1981/2005 Tmin 2006/2007
Tmin 1981/2005 Tmax2006/2007 Tmax 1981/2005
35
30
25
20
15
10
5
0
Figure 5.Températures mensuelles des
périodes 1981/2005 et 2006/2007.
120
100
40
60
20
80
0
Mois
Figure 6. Précipitations mensuelles de la compagne
2006/2007.
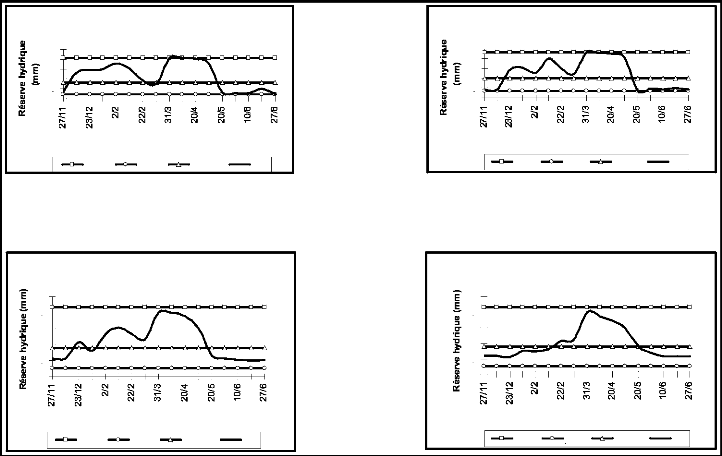
1-1-3 Etat hydrique du sol selon les horizons
Sur la figure 6 on observe une variation spatio-temporelle de
la réserve hydrique du sol. A l'échelle du temps, on constate que
pour le même horizon, la réserve hydrique présente de
très grandes fluctuations suivants les différentes dates. A
l'échelle de l'espace, on constate que pour la même date, la
réserve hydrique n'est pas similaire chez les différents horizons
(tableau 9).
Dès le semis, le sol est presque à sec. Tous les
horizons présentent des déficits hydriques qui se traduisent par
un déficit de la réserve facilement utilisable (RFU), soit;
-10,03 et -11,52 mm pour H1 et H2, et -6,76 et -5,89 mm pour H3 et H4.
Dès les premières pluies de Décembre, le
réservoir du sol commence à se remplir graduellement. Il
enregistre une réserve facilement utilisable de 8,2 mm pour H1 le 8
Décembre et atteint 8,61 mm pour H2, 14 jours après. Pour H3 et
H4, le déficit persiste plus tard et il n'est pallié qu'au mois
de Février, avec respectivement une RFU de 8,85 mm le 2 Février
pour le troisième horizon et 3,60 mm le 22 Février pour le
dernier horizon.
Mis à part un léger déficit ressentit
durant la troisième décade de février (-1,56 mm pour H1),
le réservoir du sol se remplit progressivement atteignant la
capacité au champ à la fin du mois de Mars. Le stock en eau
était constamment à son maximum sur toute la profondeur de notre
parcelle, mais à partir de la deuxième décade de Mai, il a
été noté un épuisement de la réserve en eau,
puisque sur une dizaine de jours seulement, toute la RFU a été
consommée.
A partir de la troisième décade de Mai et
jusqu'à la récolte, un déficit hydrique permanent
s'installe. Il est plus avancé pour H1 et H2, qui avoisinent le point de
flétrissement, avec une RFU respective de -11,96 et -11,45 mm,
comparés à H3 et H4 dont la RFU est estimée à -7,63
et -6,51 mm.
1-1-4 Bilan hydrique selon des stades
phénologiques En utilisant les relations suivantes:
ETR=P- ÄRU (1)
ETR : évapotranspiration réelle, P :
précipitation et ÄRU : variation de la réserve hydrique
et
ETc = ETo * Kc (2)
L'ETc est l'évapotranspiration de la culture en absence de
stress, et l'ETo est l'évapotranspiration de référence et
Kc: le coefficient cultural
40
80
70
60
50
30
40
60
80
70
50
30
Cc pF RFU RE
Cc pF RFU RE
H3 :40-60cm
H1 :0-20cm
Date
Date
40
60
80
70
50
30
40
80
70
60
50
30
Cc pF RFU RE
Cc pF RFU RE
H4: 60-80 cm
H2:20-40cm
Date
Date
Figure 7. Variation de la réserve hydrique du
sol selon les horizons.
Tableau 9. Variation de la réserve utile et de la
RFU selon les horizons (en mm).
|
Horizon (cm)
|
Date
|
|
27/11
|
08/12
|
23/12
|
05/1
|
02/2
|
12/2
|
22/2
|
04/3
|
31/3
|
10/4
|
20/4
|
10/5
|
20/5
|
30/5
|
10/6
|
20/6
|
27/6
|
|
0 - 20
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Humidité Pondérale
|
13,87
|
21,24
|
22,55
|
22,77
|
24,78
|
22,91
|
18,44
|
17,11
|
26,97
|
26,95
|
26,69
|
24,97
|
13,70
|
13,46
|
13,19
|
15,21
|
13,13
|
|
Réserve Utile
|
2,29
|
21,76
|
25,21
|
25,81
|
31,10
|
26,16
|
14,37
|
10,85
|
36,88
|
36,83
|
36,15
|
31,60
|
1,85
|
1,20
|
0,49
|
5,83
|
0,36
|
|
RFU
|
-10,03
|
9,44
|
12,89
|
13,49
|
18,78
|
13,84
|
2,05
|
-1,47
|
24,56
|
24,51
|
23,83
|
19,28
|
-10,47
|
-11,12
|
-11,83
|
-6,49
|
-11,96
|
|
20-40
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Humidité Pondérale
|
13,61
|
13,62
|
20,70
|
21,30
|
19,67
|
24,64
|
21,05
|
19,06
|
26,98
|
26,53
|
26,41
|
25,23
|
13,12
|
13,78
|
13,66
|
14,05
|
13,64
|
|
Réserve Utile
|
1,73
|
1,77
|
21,87
|
23,57
|
18,96
|
33,06
|
22,86
|
17,22
|
39,71
|
38,44
|
38,07
|
34,75
|
0,35
|
2,21
|
1,88
|
2,99
|
1,81
|
|
RFU
|
-11,52
|
-11,48
|
8,61
|
10,32
|
5,70
|
19,81
|
9,61
|
3,96
|
26,46
|
25,18
|
24,82
|
21,49
|
-12,91
|
-11,04
|
-11,37
|
-10,27
|
-11,45
|
|
40- 60
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Humidité Pondérale
|
15,18
|
15,19
|
18,93
|
17,03
|
20,92
|
22,37
|
20,80
|
19,59
|
25,75
|
25,63
|
24,86
|
22,19
|
15,87
|
15,21
|
14,92
|
14,81
|
14,86
|
|
Réserve Utile
|
5,93
|
5,97
|
16,14
|
10,95
|
21,54
|
25,48
|
21,22
|
17,92
|
34,69
|
34,35
|
32,26
|
25,00
|
7,82
|
6,00
|
5,21
|
4,92
|
5,07
|
|
RFU
|
-6,76
|
-6,73
|
3,45
|
-1,74
|
8,85
|
12,79
|
8,53
|
5,23
|
22,00
|
21,66
|
19,56
|
12,30
|
-4,87
|
-6,69
|
-7,48
|
-7,77
|
-7,63
|
|
60- 80
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Humidité Pondérale
|
15,50
|
15,47
|
15,18
|
16,66
|
16,50
|
17,05
|
18,99
|
19,29
|
25,85
|
24,84
|
23,76
|
21,91
|
17,63
|
16,10
|
15,34
|
15,27
|
15,27
|
|
Réserve Utile
|
6,80
|
6,73
|
5,93
|
9,95
|
9,51
|
11,03
|
16,30
|
17,11
|
34,95
|
32,22
|
29,27
|
24,23
|
12,60
|
8,44
|
6,37
|
6,18
|
6,18
|
|
RFU
|
-5,89
|
-5,97
|
-6,76
|
-2,74
|
-3,18
|
-1,66
|
3,60
|
4,41
|
22,26
|
19,52
|
16,58
|
11,53
|
-0,09
|
-4,25
|
-6,33
|
-6,51
|
-6,51
|
|
0 - 80
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Humidité Pondérale
|
14,54
|
16,69
|
20,73
|
20,37
|
21,79
|
23,31
|
20,10
|
18,59
|
26,57
|
26,37
|
25,99
|
24,13
|
14,23
|
14,15
|
13,92
|
14,69
|
13,88
|
|
Réserve Utile
|
16,76
|
36,22
|
69,15
|
70,29
|
81,11
|
95,74
|
74,75
|
63,09
|
146,23
|
141,83
|
135,76
|
115,57
|
22,62
|
17,86
|
13,95
|
19,92
|
13,42
|
|
RFU
|
-28,31
|
-8,77
|
24,95
|
22,07
|
33,33
|
46,44
|
20,18
|
7,72
|
73,02
|
71,35
|
68,22
|
53,07
|
-28,25
|
-28,85
|
-30,68
|
-24,53
|
-31,03
|
Tableau 10. Précipitation, ETR et variation de la
réserve utile (en mm).
|
27/11
|
8/12
|
23/12
|
5/1
|
2/2
|
12/2
|
22/2
|
4/3
|
31/3
|
10/4
|
20/4
|
10/5
|
20/5
|
30/5
|
10/6
|
20/6
|
27/6
|
Total
|
|
|
|
|
|
|
|
-
|
-
|
|
|
|
-
|
-
|
|
|
|
|
|
|
ÄRU
|
|
19,46
|
32,93
|
1,13
|
10,82
|
14,63
|
20,99
|
11,65
|
83,14
|
-4,40
|
-6,08
|
20,19
|
92,95
|
-4,76
|
-3,91
|
5,97
|
-6,51
|
-3,34
|
|
Pluies
|
|
19,80
|
34,00
|
8,00
|
11,50
|
20,20
|
0,00
|
1,00
|
95,60
|
37,30
|
64,90
|
32,90
|
0,30
|
1,00
|
1,00
|
16,20
|
0,00
|
343,70
|

ETR
0,34
1,07
6,87
0,68
5,57
20,99
12,65
12,46
41,70
70,98
53,09
93,25
5,76
4,91
10,23
6,51
347,04
43
S'il n'y a pas de stress, l'ETR sera égale à l'ETc.
Mais si il y'a un stress, l'ETc est ajustée par l'introduction du
coefficient de stress; Ks, suivant la relation (3):
ETcaj=ETo*Kc*Ks (3)
L'évapotranspiration réelle cumulée
durant tout le cycle cultural est de 347,04 mm pour une pluviométrie
totale de 343,70 mm, soit un déficit sur la réserve hydrique de
3,34 mm (tableau 10). L'évolution de l'ETR,l'ETo et l'ETc est
liée aux stades phénologiques et diffère tout au long du
profil (0-80 cm) pour chacun des paramètres cités.
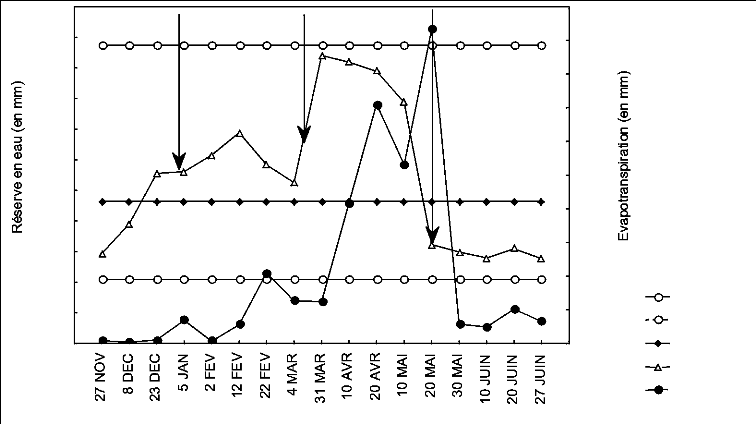
1-1-4-1 Semis - Levée
Cette phase correspond à l'installation de la culture,
durant laquelle le taux de la couverture végétale est très
faible. Elle a duré en moyenne 36,25 jours, marquée par un
léger déficit hydrique (figure 8).
L'évapotranspiration réelle (ETR) pour ce stade
est relativement faible soit ; 6,16 mm, alors que l'évapotranspiration
potentielle de la culture (ETc) est de 12,25 mm, (tableau 11), soit une
différence de 6,08 mm. L'ajustement de l'ETc par l'utilisation de la
relation (3) donne un coefficient de stress Ks1 = 0,5, ceci signifie que
l'évapotranspiration réelle était déficitaire de
près de 50% de celle prédite pour cette phase.
Tableau 11. Bilan hydrique selon les stades
phénologiques.
|
Paramètre
|
Semis
-
Levée
|
Levée
-
Montaison
|
Montaison
-
Epiaison
|
Epiaison
-
Maturité
|
Cycle
total
|
|
Durée (j)
|
36
|
70
|
70
|
32
|
208
|
|
P (mm)
|
61,80
|
84,20
|
179,50
|
18,20
|
343,70
|
|
ETR (mm)
|
6,16
|
45,70
|
268,36
|
26,83
|
347,04
|
|
ETo (mm)
|
35,00
|
115,20
|
200,40
|
242,10
|
592,70
|
|
Kc
|
0,35
|
0,75
|
1,2
|
0,73
|
0,87
|
|
ETc (mm)
|
12,25
|
86,4
|
240,48
|
176,73
|
515,86
|
|
Ks
|
0,50
|
0,53
|
1,12
|
0,15
|
0,67
|
1-1-4-2 Levée - Début montaison
Cette phase se caractérise par l'augmentation du taux
de la couverture végétale du sol, un développement
considérable du système racinaire et une sensible hausse des
besoins en eau de la culture. Ce stade s'étalé sur 70 jours. Il
enregistre un cumul d'ETR et de d'ETc de 45,70 et 86,40 mm, respectivement. Le
coefficient de stress pour ce Ks2 = 0,53. Ce stress est enregistré
durant la période du 12 Février au 4 Mars, correspondant au stade
plein tallage à début montaison. Ceci malgré que la
réserve hydrique sur l'ensemble du profil (0 - 80) cm était au
dessus de la RFU (figure 7).
280
260
240
220
200
320
300
180
160
140
120
100
Semis-Levée
Levée-Montaison Montaison-Epiaison
Epiaison-Maturité
40
20
90
80
70
60
50
30
0
100
10
CC pF RFU RE ETR
Figure 8. Evapotranspiration réelle et variation
de la réserve hydrique sur le profil 0 -80 cm.
On note un épuisement progressif de la réserve
hydrique (figure 7) suivant les horizons H1, H2, puis H3. Soit une RFU
respective de: -1,47 ; 3,96 et 5,23 mm durant la première semaine de
Mars. Ceci serait probablement la conséquence du manque de
précipitations, puisqu' il n'a été enregistré qu'un
mm du 10 Février au 07 Mars, d'une part, et d'autre part par
l'élévation brusque des températures maximales avoisinant
en moyenne 20,41 °C durant la période du 28 Février au 06
Mars.
1-1 -4-3 Début montaison - Epiaison
Cette phase a durée 70 jours. Il a été
enregistré 179,50 mm de pluie, représentant l'équivalent
de 52% de tout le cycle cultural. Elle totalise à elle seule, 77% de
l'ETR du cycle.
Durant les 60 premiers jours, la RFU est positive avec 73,02
et 53,07 mm respectivement pour les dates : 31 Mars et le 10 Mai. Ceci laisse
supposer que ces deux mois se sont déroulés dans de bonnes
conditions d'alimentation hydrique du sol. En revanche durant la
deuxième décade de Mai, on enregistre seulement 0,3 mm de pluie
pour une ETR estimée à 93,35 mm, représentant 1/3 de l'ETR
de tout le stade. Ceci a pour conséquence, l'épuisement de la
réserve hydrique du sol, puisqu'il a été noté, pour
la même décade un ÄRU égale à 92,25 mm et une
chute de la RFU à -28,25 mm au 20 Mai.
L'ETc calculée pour ce stade est estimée
à 240,48 mm pour une ETR de 268.36 mm, soit un Ks3 = 1,12. Mais, en
calculant l'ETc du début de ce stade jusqu'au 10 Mai, on obtient une ETc
= 176,28 mm, qui est très proche de l'ETR = 174,53 mm, avec un Ks3' =
0,99. Du 10 au 20 mai l'ETc est égale à 64,2 mm pour une ETR
égale à 174,53 mm soit, un Ks3» = 1,46. La différence
entre l'ETc et l'ETR pour seulement cette décade, représente
près de 46%; ceci est probablement dû à une sous estimation
du coefficient culturale (Kc) pour cette décade (tableau 12) où
il a été observé une élévation significative
des températures, qui passent d'une moyenne de près de 13 °C
à près de 18 °C et des maxima atteignant près de 24
°C. Cette élévation dans les températures a peut
être accentué l'évapotranspiration réelle de cette
décade.
Tableau 12. Bilan hydrique du stade; début
montaison à l'épiaison.
|
Paramètre
|
du 12Mars au 10 Mai
|
du 10Mai
au 20 Mai
|
du 12Mars
au 21 Mai
|
|
P (mm)
|
179,2
|
0,3
|
179,5
|
|
ETR(mm)
|
174,53
|
93,83
|
268,36
|
|
ETo (mm)
|
146,9
|
53,5
|
200,4
|
|
KC
|
1,2
|
1,2
|
1,2
|
|
ETc(mm)
|
176,28
|
64,2
|
240,48
|
|
Ks3
|
0,99
|
1,46
|
1,12
|
1-1-4-4 Epiaison - Maturité
Ce dernier stade de développement, qui va de
l'épiaison jusqu'à la maturation, a duré 32 jours. Il se
caractérise par une forte ETc (176,73 mm) et un faible ETR (26,83 mm).
En effet, la conjugaison de la faible pluviométrie (18,20 mm) et
l'épuisement de la RFU (allant de -28,25 à -31,03 mm du 20 Mai
jusqu'à la récolte) ont fait que ce stade a été
caractérisé par un stress hydrique intense (Ks = 0,15).
|
|


