III - 2.4 Le croissement
Dans un algorithme génétique, des parties des
individus sélectionnés (parents) sont échangées par
croisement. Le croisement peut être effectué sur un ou plusieurs
parents pour former un ou plusieurs enfants (ou descendants). Il existe,
là aussi, de nombreuses méthodes de croisement.
Nous présentons ici les croisements classiques, qui
sont le croisement en un point, le croisement en deux points et le croisement
uniforme, il y à d'autres méthodes de croisement
appelées le croisement diagonal et le croisement de bloc. Ces derniers
opérateurs sont bien adaptés à la transmission des
propriétés topologiques entre les parents et les descendants
[25].
III - 2.4.1 Croisement en un point
On choisit au hasard un point de croisement, pour chaque
couple figure III-2. Notons que le croisement s'effectue
directement au niveau binaire, et non pas au niveau des gènes. Un
chromosome peut donc être coupé au milieu d'un
gène.
2 parents 2 enfants

10010
1001101011
10000
011010111
100100
001101
100100 10101001001
001101 10011010111
Figure III-2 : Principe de
croissement en un point
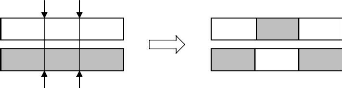
III - 2.4.2 Croisement en un et deux points
On choisit au hasard deux points de croisement figure III-3.
Par la suit, nous avons utilisé cet opérateur car il est
généralement considéré comme plus efficace que la
précédent. Néanmoins nous n'avons pas
constaté de différence notable dans la convergence de
l'algorithme.
Notons que d'autre formes de croisement
existent, du croisement en K points jusqu'au cas limite du
croisement uniforme.
2 parents 2 enfants
10101
001101
100100
001001
010111
10011
100100 10101 001001
001101 10011 010111
Figure III-3 : Principe de
croissement en deux points
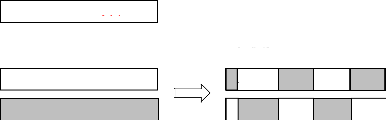
III - 2.4.3 Croisement uniforme
Le croisement binaire utilise un masque (en binaire) de
même longueur que les parents. Chaque 1 contenu dans le masque
déclenche de l'inversion du gène visé
avec celui de l'autre parent. Voir le figure III-4 [25] :
0 0 0 1 1 1
Masque
2 enfants
0
0 0 1 0 0 1
0 010 0
0 1 0 1 1 1
011 0 1
1
|
1 0 0
|
0 0
|
0 1
|
1 1
|
0 0
|
|
2 parents
|
|
|
|
1 0 0
|
1 0
|
0 1
|
0 1
|
0 1
|
|
|
|
|
|
|
0 0 1
|
1 0
|
1 1
|
0 0
|
1 1
|
|
100
|
01 00
|
1
|
110 1
|
|
|
|
|
|
100
|
1 10 1
|
0
|
1 1
00
|
Figure III-4 : Croisement
uniforme
III - 2.5 Mutation
Nous définissons une mutation comme étant
l'inversion d'un bit dans un chromosome. Cela
revient à modifier aléatoirement la valeur d'un
paramètre du dispositif. Les mutations jouent le rôle de bruit et
empêchent l'évolution de se figer. Elles
permettent d'assurer une recherche aussi bien globale que
locale, selon le poids et le nombre des bits mutés. De plus, elles
garantissent mathématiquement que l'optimum global peut
être atteint.
1 0 0 1 0 0
1 0 0 1 0 0
0 1 0 1 0 0 1 0 0 1
0 0 1 0 1 0 0 1 0 0 1
Une mutation
1 1

Figure III-5 : Opérateur
de mutation
Début
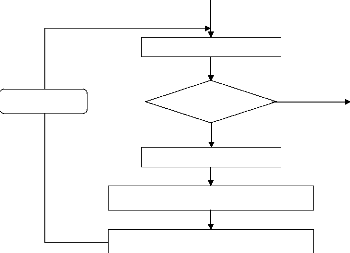
III - 2.6 Organigramme de la procédure
génétique
L'organigramme fonctionnel de la figure III-6
illustre la structure de l'algorithme génétique
standard. Nous détaillerons les diverses phases qui le constituent et
présenterons les mécanismes associés à chacune
d'entre elles dans les sections suivantes [26].

Population initiale
T=T+1
Recombinaison : Croisement
Nouvelle Population
Non
Evaluation
Terminer
Sélection
Oui
Résultat
Figure III-6 : Organigramme d'un
algorithme génétique.
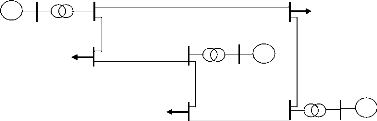
III - 2.7 Application de l'AG à la
répartition économique des puissances
Nous appliquons ici l'algorithme
génétique de base étape par étape à la
répartition économique des puissances sur un simple réseau
test constitué de 9 jeux de barres, 6 lignes électriques, 3
générateurs, 3 transformateurs et 3 charges.
G
G
G
Figure III-7 : Schéma
unifilaire de réseau électrique
Le tableau III-2 montre les données techniques et
économiques des trois générateurs interconnectés.
La puissance de base utilisée est de 100 MVA.
|
°
N
|
Pmin ( Pu )
|
Pmax ( Pu )
|
($ / h )
|
( $ / Mwhr)
|
( Mw2 hr)
$ /
|
|
1
|
0.30
|
1.8
|
105.0
|
2.45
|
0.01
|
|
2
|
0.15
|
0.9
|
44.1
|
3.51
|
0.01
|
|
3
|
0.40
|
1.9
|
40.6
|
3.89
|
0.01
|
Tableau III-2 : Ensemble des
paramètres des puissances actives générées
PGi
Le problème de la REP consiste à trouver le minimum
de la fonction objective. Chaque puissance active générée
PGi est limitée par une limite inférieure PGi min et une
limite supérieure PGi max . Puisque la
fonction objective est bornée supérieurement, on va
choisir une fonction sélective à maximiser de la forme suivante
[22] :
|
fitness
|
? F - F x
( ) si F x
( ) <
max
= ?
?0 sinon
|
Fmax
|
III - 2.7.1 Codage des chromosomes et le
décodage
La première étape consiste à coder les
variables PGi sous forme de chromosome. Pour sa
simplicité et sa commodité, le codage binaire est utilisé
dans cet exemple. Avec le codage binaire, les puissances actives
générées du réseau test(PG 1 ,
P G2 , PG3) vont
être codées comme une chaîne de « 0
» et « 1 »
avec,
respectivement, des longueurs(L1 , L
2 , L 3 ) (peuvent être
différentes).
Chaque paramètre PGi a une limite
supérieure PGi max = B et une limite inférieure
PGi min = A. Le choix de L1,L 2
et L3 pour les paramètres est sujet de la
résolution spécifiée par l'utilisateur
dans l'espace de la
recherche. Avec le codage binaire, la relation entre la longueur
de bit Li et la résolution correspondante Ri est donnée par :
Donc, l'ensemble PGi peut
être transformé en une chaîne binaire (chromosome) avec une
longueur
? Li et puis l'espace de
recherche est exploré. Il est à noter que chaque chromosome
présente une solution possible du problème. Par exemple, supposer
le domaine des paramètres de (PG 1 , P
G2 , PG3) qui est
présenté dans le Tableau 1. Si la résolution R
1 , R 2, R3 est
spécifiée comme 0.1, 0.05, 0.1 d'après
l'équation (7) on aura L1, L
2 et L3 = (4,4,4) . Alors
l'ensemble de paramètres (PG 1 ,
P G2 , PG3 ) peut
être codé selon le tableau III-3.
|
N°
|
P1 (Pu)
|
Code
|
P2 (Pu)
|
Code
|
P3 (Pu)
|
Code
|
|
1
|
0.3
|
0000
|
0.15
|
0000
|
0.4
|
0000
|
|
2
|
0.4
|
0001
|
0.20
|
0001
|
0.5
|
0001
|
|
3
|
0.5
|
0010
|
0.25
|
0010
|
0.6
|
0010
|
|
4
|
0.6
|
0011
|
0.30
|
0011
|
0.7
|
0011
|
|
5
|
0.7
|
0100
|
0.35
|
0100
|
0.8
|
0100
|
|
6
|
0.8
|
0101
|
0.40
|
0101
|
0.9
|
0101
|
|
7
|
0.9
|
0110
|
0.45
|
0110
|
1.0
|
0110
|
|
8
|
1.0
|
0111
|
0.50
|
0111
|
1.1
|
0111
|
|
9
|
1.1
|
1000
|
0.55
|
1000
|
1.2
|
1000
|
|
10
|
1.2
|
1001
|
0.60
|
1001
|
1.3
|
1001
|
|
11
|
1.3
|
1010
|
0.65
|
1010
|
1.4
|
1010
|
|
12
|
1.4
|
1011
|
0.70
|
1011
|
1.5
|
1011
|
|
13
|
1.5
|
1100
|
0.75
|
1100
|
1.6
|
1100
|
|
14
|
1.6
|
1101
|
0.80
|
1101
|
1.7
|
1101
|
|
15
|
1.7
|
1110
|
0.85
|
1110
|
1.8
|
1110
|
|
16
|
1.8
|
1111
|
0.90
|
1111
|
1.9
|
1111
|
Tableau III-3 : Codage de
l'ensemble des paramètres de PGi
Pour construire un codage multi-paramétrié, on
peut tout simplement concaténer autant de codage d'un
seul paramètre qu'il est nécessaire. Le
chromosome correspond à l'ensemble de paramètres
(1.7, 0.30, 1.1) est alors la chaîne de caractères binaire
suivante 111000110111. Le décodage est la procédure inverse
| 

