Recherche de facteurs explicatifs des écarts à la relation négative rendement - teneur en protéines chez le blé tendre (Triticum aestivum L. )( Télécharger le fichier original )par Fatma Ben Cheikh Institut des sciences et industries du vivant et de l'environnement, dit AgroParisTech - Master 2 recherche 2012 |

Objectifsï Calcul du GPD pour caractériser la variabilité interannuelle du climat et multilocale affectant la relation rendement - teneur en protéines ï A l'instar du travail fait par Mattieu Bogard (2011) dans le cadre d'une thèse qui porte sur l'explication du GPD via la variabilité génétique, vérifier s'il est possible d'expliquer les variations interannuelles et multilocales de GPD par un surplus d'absorption d'azote en post-floraison. ï Rechercher les variables agroclimatiques corrélées au GPD et/ou au surplus d'absorption post-floraison. Analyse bibliographique1. Fonctionnement azoté et élaboration de rendement et taux de protéines 1.1. Métabolisme azotée dans la planteL'azote est un élément majeur de la nutrition chez le blé. L'introduction des variétés « semi-naines » a permis d'accroître les quantités d'azote valorisables par un couvert de blé (Barraclough et al., 2010). L'azote agit sur la croissance de l'appareil végétatif, en contrôlant la taille des feuilles et en jouant sur le devenir des talles. Il influence également fortement le nombre d'épillets fertiles. Lorsque le statut azoté de la culture passe au-dessous du seuil défini comme l'état permettant la croissance maximale de la culture, celle -ci est en carence. Cette carence azotée agit sur les fonctions de croissance en réduisant l'accumulation de biomasse, l'indice foliaire, son expansion ou en accélérant le processus de sénescence. La contrainte azotée peut également affecter l'efficience de conversion de l'énergie lumineuse (Jeuffroy et Recous, 1999). L'azote est absorbé par les racines sous ses deux formes ; nitrique (NO3-) et ammoniacale (NH4+). L'absorption se fait grâce à des transporteurs au niveau de la racine (NRT: nitrate transporter et AMT: ammonium transporter). Dans la plante l'ion nitrate prend deux chemins Il subit une réduction dans les feuilles pour enfin aboutir à l'ion NH4+. Cependant, dans le cas de faible besoin en azote il est stocké dans les vacuoles pour constituer un pool de réserve. (Gate, 1995). De l'autre coté l'ammonium est assimilé dans les racines. Les produits de l'assimilation de l'azote sont des acides organiques et acides aminés. (Morot-Gaudry, 1997) L'assimilation de l'azote se fait à l'aide de deux enzymes le GS (glutamine synthétase) et la GOGAT (glutamate synthase). Ces deux enzymes intègrent l'ammonium dans une molécule organique qui sera à l'origine de la formation de deux acides aminés: la glutamine et le glutamate. Des protéines seront par la suite formées grâce à des acides aminés dérivant du glutamine et le glutamate (Masclaux et al., 2001 cité par Ben Slimane, 2010). L'élaboration du rendement dépend de l'absorption azotée. Cette absorption se fait selon la disponibilité de l'élément dans le sol et est fonction des besoins de la plante. En effet, les besoins de la culture varient selon les stades de croissance. On a montré que les plus forts besoins sont entre la phase d'épi 1 cm et la floraison. Un déficit azoté pendant cette période affecte le rendement. Au cours de la montaison, l'azote agit sur la montée à épis des talles herbacées, la croissance des feuilles et l'élongation des entre-noeuds. De plus, une carence en azote dans la phase de montaison altère le nombre de grains (Jeuffroy et Recous, 1999). L'accumulation de biomasse dans les grains est le résultat de la confrontation entre la demande des grains en biomasse et l'offre en assimilats carbonés au niveau de la culture. La quantité de biomasse disponible pour le remplissage des grains est dépendante de la production de biomasse par la culture après la floraison (source majoritaire des assimilats carbonés pour les grains) et d'une fraction fixe de la biomasse à floraison, correspondant à la remobilisation des sucres solubles stockés dans les tiges avant ce stade (O'Leary et al., 1985 cité par Barbottin, 2004). La nutrition azotée intervient donc dès le tallage avec des effets dominants à partir du stade épi 1 cm jusqu'à la floraison sur les composantes du rendement : nombre d'épis/m² et nombre de grain (Gate, 1995).
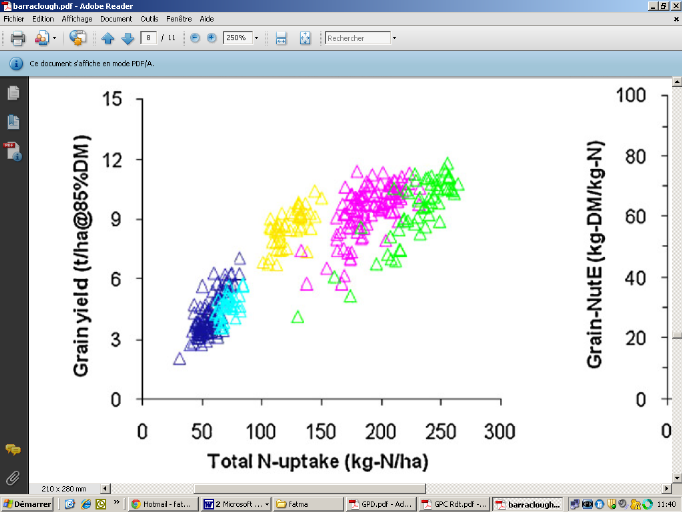
Figure 1: Relation entre le rendement (t/ha) et quantité d'azote absorbée (kg N /ha) (les différentes couleurs des triangles expliquent les doses d'azote) (Barraclough et al., 2010)
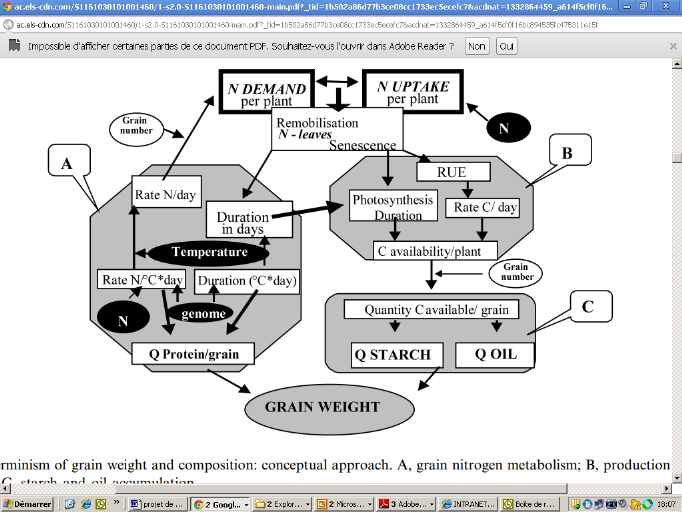
L'azote assimilé disponible au niveau des grains est essentiellement utilisé pour la synthèse des protéines qui seront accumulées et stockées dans l'endosperme. (Triboi et Triboi-Blondel, 2002) L'azote est stocké dans le grain sous forme de protéines. Le stockage se fait essentiellement au cours de la période post-floraison grâce à la remobilisation de l'azote des parties végétatives en cours de sénescence. (Shewry, 2007). Il faut noter que l'accumulation d'azote dépend de la disponibilité en azote au niveau de la plante. (Barbottin, 2004) On a montré que même un faible apport en azote dans la phase post-floraison affecte positivement la teneur en protéine (Gate, 1995). Trois modalités de fertilisation azotée ont une influence sur la teneur en protéines : - la dose apportée qui avec laquelle la teneur en protéine augmente. - la forme d'azote utilisée où on constate que la forme nitrate permet une augmentation du taux de protéines comparée aux formes ammoniaco-uréiques. - le fractionnement des apports, surtout en appliquant un apport tardif qui permet d'augmenter la teneur en protéines les apports tardifs grâces à une meilleure utilisation de l'azote par le blé, permettent d'augmenter la teneur en protéines (Anonyme 1, 2011). La figure (2) est un diagramme séparant les deux mécanismes azoté et carboné pour mieux comprendre leur interaction et leur impact sur l'établissement de rendement et la teneur en protéines.
Figure 2: Déterminisme du poids des grains et sa composition: A, métabolisme azoté dans le grain; B: production des assimilats de carbone; C: accumulation de l'amidon (Triboi et Triboi-Blondel, 2002) 2. Relation entre le rendement et la teneur en protéines
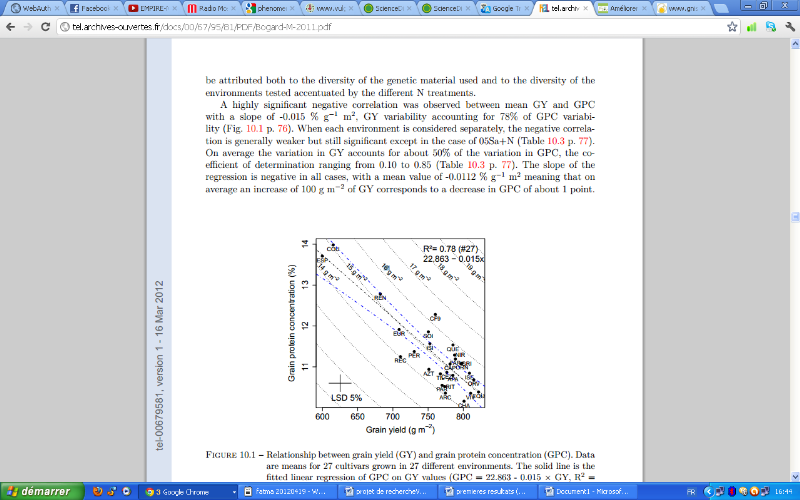
Les deux critères : rendement en grain et la teneur en protéines étaient l'objet de plusieurs recherches. Ainsi la relation entre ces deux variables était bien étudiée. La majorité des études montrent qu'entre le rendement et la teneur en protéines la relation est plutôt négative. (Monaghan et al, 2001, Oury et Godin, 2007, LeGouis et al.,2010). Ces résultats ont été confirmés récemment par l'étude de Bogard (2011). Il a trouvé une relation négative très significative entre les deux critères ( Figure 3)
Figure 3: Relation entre le rendement en grain et la teneur en protéines pour 27 variétés dans 27 milieux différents (Bogard, 2011)
Une première étude était faite dans le cadre d'expliquer la relation entre le rendement et la teneur en protéines par rapport aux variétés. La comparaison était faite entre des variétés anciennes et des variétés récentes dites « performantes ». Les résultats étaient que les variétés anciennes ont un taux de protéines élevé mais à faible rendement. Tandis que les variétés récentes affichent un bon rendement contre une teneur faible en protéines (Barraclough et al., 2010). Ceci met en évidence la relation négative qui existe entre les deux variables. Dans le but d'expliquer le paradoxe de la relation, certains auteurs Pepe et Robert, 1975 ; Miezen et al., 1977, McNeal et Berg, 1978 (cité par Bogard, 2011) ont trouvé que c'est le facteur génétique est responsable de cette relation. Ils ont proposé que la nature de la relation soit due essentiellement aux facteurs génétiques par la présence des gènes à effet pléiotropes affectant les deux caractères ou bien par la présence des gènes à effet antagoniste fortement liés génétiquement. A côté du facteur génétique, une forte hypothèse concernant la compétition entre les métabolismes azotés et carbonés peut expliquer cette relation négative (bhatia et rabson, 1976 cité par Bogard 2011). On a montré qu'il existe une étroite interaction entre ces deux métabolismes. L'azote est remobilisé des organes végétatifs vers les grains via le processus de sénescence : par conséquent, la perte d'azote dans les organes végétatifs contribue à réduire l'indice foliaire et l'efficience de conversion du rayonnement en biomasse. Le remplissage des grains en biomasse, qui dépend de l'interception lumineuse pilotée par l'indice foliaire et de son efficience de conversion, est donc directement impacté par cette remobilisation (Jeuffroy et al., 2000 ). La nature de la relation entre le rendement et la teneur en protéines résulte essentiellement des interconnexions entre métabolismes carbonés et azotés (Bogard, 2011). Ces deux variables résultent de l'intégration de processus en rapport à la fois avec les métabolismes du carbone et de l'azote. Le rapport C/N du grain étant largement en faveur du carbone, le rendement en grains dépend en premier lieu des facteurs déterminant l'assimilation photosynthétique, le stockage et la remobilisation des hydrates de carbone. Pour la teneur en protéines, cela dépend fortement de la quantité d'azote absorbée et remobilisée vers le grain (Bogard, 2011). La teneur en protéines des grains dépend de la richesse de la plante en acides aminés durant la période post-floraison. Les acides aminés, pendant la méiose, permettent la formation des protéines de structure et métabolique, qui, en phase de remplissage deviennent des protéines de réserve. (Martre et al., 2003 cité par Bertheloot, (2009). Le développement des grains nécessite la réalisation d'un mécanisme dont le processus permet le recyclage des protéines des organes végétatifs via leur dégradation en acides aminés et leur transfert vers les organes en croissance (les grains en post-floraison): c'est la remobilisation (Hirel et al., 2007 cité par Ben Slimane, 2010). Un nombre élevé de grain accélère la remobilisation de l'azote engendrant une accélération de la sénescence pendant le remplissage et qui par conséquent affecte la photosynthèse. (Bogard et al., 2011). Selon Przulj et Momcilovic (2001) la concentration en protéines dans le grain ne dépend pas seulement de la quantité d'azote dans le grain mais aussi de la concentration en carbohydrates. On a montré que cet effet antagoniste entre les deux caractères de la production est essentiellement dû à un phénomène de dilution de l'azote absorbé au profit du carbone Slafer et al. 1990 cité par Monaghan et al (2001) ont montré que l'augmentation de la teneur en protéines, peut se faire par une accumulation plus importante de l'azote absorbé en préfloraison combinée avec une forte remobilisation de l'azote des parties végétatives vers les grains. 3. Concept de grain protein deviation « GPD » Le GPD a été étudié pour la première fois par Mongham et al (2001) en Angleterre dans le but de mesurer la capacité des variétés de blé tendre d'atteindre un taux de protéines meilleur que prévu par rapport un rendement donné. Pour évaluer la relation négative reliant le rendement et la teneur en protéine, on a proposé d'utiliser la déviation à partir de la régression linéaire entre le rendement et la teneur en protéine ; c'est ce qu'on appelle « la déviation grain protéine » : « Grain Protein deviation » (Anonyme2, Monaghan et al., 2001, Oury et Goudin, 2007, LeGouis et al, 2010, ...). Le GPD étant définie comme étant le « résidus standardisés » de la régression linéaire du rendement en grain (t/ha) et la teneur en protéines (%) (Oury et Godin, 2007 ; Monghan et al., 2001 ). Oury et Goudin (2007) ont proposé d'utiliser une sorte d'algorithme d'ajustement dans le but d'éliminer les outliers et évaluer la vraie position de la régression linéaire pour un ensemble de données. L'algorithme fonctionne en éliminant tous les points dont les résidus sont supérieurs à |1.96|. L'itération est stoppée quand on a plus de valeur en dessus de la valeur seuil Dans leur étude Oury et Godin (2007) ont utilisé la formule (1) pour calculer les résidus de la régression : GPD=Sd= ri / ^s*v (1-hi) (1); ^s est la déviation résiduelle standard ; ^s=v((1/n-p)*?ri²) Avec : ri= résidus des observations, p=2, n=nombre des observations, hi=la valeur de la diagonale de la matrice. En effet, étant donné que les GPD sont des résidus standardisés donc, par définition, dans la condition d'une distribution normale, la moyenne de GPD est nulle et 95% des valeurs de GPD standardisées sont comprises entre -1.96 et +1.96. Cette dernière valeur correspond au quantile d'ordre 2.5% pour P=0.975. L'une des méthodes utilisées pour obtenir le GPD est d'utiliser un algorithme pour éliminer tous les points qui ont des résidus supérieur à |1.96| l'itération est stoppée quand on a plus de valeur en dessus de la valeur seuil. Le but de cette méthode est de neutraliser l'effet des données aberrantes. On aura donc une régression linéaire précise et qui représente tous les points (Oury et Godin, 2007). Afin de décomposer le GPD et d'identifier les caractères associés, la méthode d'analyse a consisté à rechercher les corrélations permettant d'établir un lien entre ce caractère et d'autres variables. Pour cela, les coefficients de corrélation des indicateurs statistiques jugeant de la force de l'association entre deux variables ont été analysés (Bogard et al., 2011). Monaghan et al. (2001) ont utilisé la méthode de « step-wise multiple régression » qui consiste à ajouter des variables pour voir leur contribution dans l'explication du GPD. Ils ont constaté que l'accumulation de l'azote post-floraison et la quantité d'azote remobilisée sont les variables les plus importantes qui ont une influence sur le GPD.
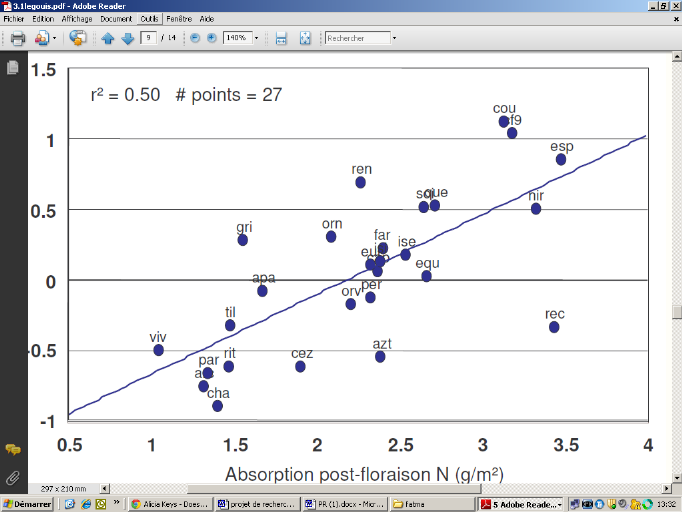
Le Gouis et al ont montré qu'il y a une corrélation entre la quantité d'azote remobilisé et la quantité d'azote absorbé post-floraison avec l'écart à la relation rendement teneur en protéines. Cependant, la corrélation n'est pas significative entre la quantité d'azote remobilisée et le GPD. La corrélation est par contre significative avec la quantité d'azote absorbé en post-floraison qui explique environ 50 % du GPD (Le Gouis et al., 2010).
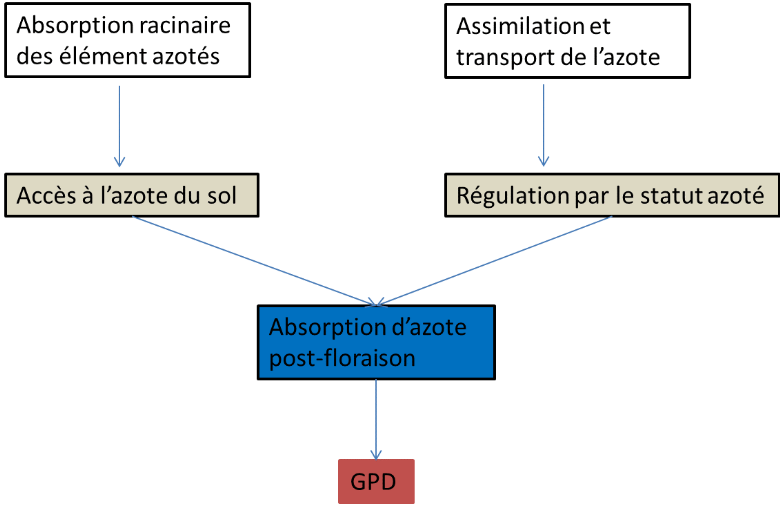
Figure 4: Processus physiologique affectant le GPD chez le blé (Bogard, 2011) Bogard (2011) en étudiant la relation entre GPD et l'azote absorbé pendant la période post-floraison a trouvé que ces deux variables sont très bien corrélés indépendamment de la date de floraison (figure 6). Monaghan et al (2001) ont trouvé que l'explication de l'importance du PANU dans la détermination du GPD est liée à la différence de la répartition de l'azote accumulé en floraison et après floraison L'absorption post- floraison, bien que relativement faible, est une source significative d'azote pendant la phase de remplissage (Le Gouis et al., 2010) mais elle est très variable. Elle dépend du site, du type de sol notamment de sa réserve d'azote disponible à la plante ainsi que de la réserve utile. Sur plantes attaquées par une maladie foliaire on peut observer l'importance de l'absorption azotée post-foliaire. Bancal et al., (2008) ont observé que l'accumulation d'azote dans les parties aériennes s'arrête plus tard dans des parcelles traitées aux fongicides que dans celles où les plantes sont attaquées par une maladie fongique. De façon assez générale, le bilan d'azote entre floraison et maturité montre que l'absorption de l'azote post-floraison est significativement réduite chez les plantes malades avec un effet significatif de l'année, donc de la sévérité des maladies et souvent aussi de la fertilisation azotée. La diminution de l'absorption semble donc fonction de la précocité et de la sévérité de l'infection. Leitch et Jenkings (1995) ont observé une réduction de 11% de l'absorption post-floraison dans le cas d'une infestation par la Septoriose foliaire, mais sans effet de la fertilisation azotée (Ben Slimane, 2010) D'autre part l'étude des variétés « stay-green » de sorgho met en évidence l'importance de l'absorption azotée post-floraison. Les « stay-green » sont des variétés qui ont une activité photosynthétique plus longue même sous des conditions hydriques limitantes. Ces variétés sont caractérisées par une concentration d'azote dans les différents compartiments toujours élevée. En effet, cette expression de « stay-green » est la conséquence de l'équilibre entre la demande en azote des grains et la réserve de l'azote durant le remplissage (Borell et Hammer, 2000). Borell et Hammer (2000), pour une étude sur le sorgho ont montré que la concentration de l'azote dans les différents compartiments de la plante est toujours plus élevée que chez les autres variétés. GPD 251657728 Absorption post-floraison N (g/m²) 251656704 Figure 5: Relation entre absorption post-foliaire de l'azote et le GPD (Anonyme 3) L'azote remobilisé est définie comme étant la quantité d'azote mesurée en floraison et qui n'a pas été récupérée dans les pailles à la récolte Pask et al (2011). L'étude de cette portion d'azote est intéressante puisque elle constitue la majeure source de la présence de l'azote dans le grain lors du remplissage. Elle représente 60 à 90% de l'azote des grains (Pask et al., 2011 ; Barbottin, 2005). La remobilisation de l'azote dépend fortement de la quantité de l'azote absorbé par la culture pendant la phase de floraison. En effet ; la quantité d'azote remobilisée est d'autant plus importante que la quantité d'azote absorbé en floraison est importante. Toutefois, la remobilisation de l'azote des parties végétatives vers les grains dépend de plusieurs facteurs environnementaux (stress hydrique et stress thermique) au cours de la période de remplissage Elle dépend aussi de l'état sanitaire des plantes : en cas de maladies foliaires, la quantité d'azote remobilisée sera fortement affectée. De plus on a indiqué que la remobilisation de l'azote est indépendante du facteur génétique dans le cas où il n'y a aucun facteur environnemental limitant (Barbottin et al., 2005). Une étude sur l'orge a montré les mêmes résultats obtenus sur le blé tendre et on a montré le rôle que joue la remobilisation de l'azote sur le rendement. En effet, le rendement est d'autant plus élevée que la quantité d'azote remobilisée est importante. (Przulj et Momcilovic, 2001). Ces études mettent en avant l'importance de la remobilisation de l'azote comme un caractère pouvant expliquer le GPD. |
|