1.2.7.1 La double paroi cellulaire décrite par un
empilement multicouche de sous couches à
fibres parallèles.
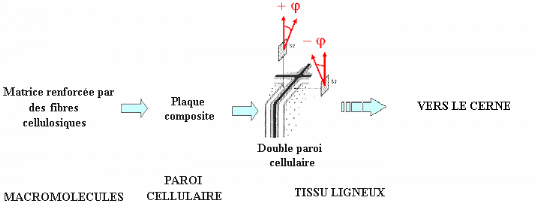
Figure 1.5 Transition du comportement mécanique
d'une sous couche à celui de la double paroi par la création d'un
multicouche à plis anti symétriques (tiré et adapté
de Salmèn et al, 1986). La notation ö représente ici l'AMF
propre à
la sous couche S2.
Ce type de modélisation (Carlsson et al,
1986, Frey-Wyssling, 1968 dans Bodig, 1982, Barrett, 1973, Gillis,
1972, Cave, 1975,1976, Mark, 1980, Koponen et al, 1989, 1991,
Norimoto, 1986, Salmén, 1986), visant à décrire le
comportement élastique de la double paroi cellulaire, fait appel
à la théorie classique des stratifiés (Tsai et
al, 1980, Daviaud et al, 1983, Guitard et al, 1987).
Cette deuxième étape de la
modélisation intégrative micro-macro, assure la transition
mécanique de l'échelle de la sous couche à celle de la
double cloison. Elle permet notamment d'exprimer les rigidités de
membrane (sous l'hypothèse de contraintes planes) et conduit, in fine,
à des estimations des modules bipariétaux longitudinal et
transverses (double paroi).
Les modèles multicouches les plus simples
définissent ainsi ce même passage par un assemblage de deux
sous couches S2 (à fibres parallèles) parfaitement
collées. D'autres, plus complexes et certainement cytologiquement
plus réalistes, autorisent diverses distinctions
(épaisseurs, composition chimique, AMF) entre les
différentes sous couches envisagées.
Par l'empilement de sous couches (plis) à inclinaison de
fibres de renforts antisymétrique, la double cloison est décrite
par un stratifié non équilibré. Par conséquent, on
notera la présence
de termes de couplages non nuls entre les rigidités de
membrane et les moments de flexion torsion.
Nous reviendrons sur les expressions des
différentes caractéristiques élastiques des
multicouches pariétaux au cours des chapitres suivants (chapitres 2 et
3).
19
Inventaire des facteurs essentiels de la variabilité intra
arbre des propriétés mécaniques des tissus ligneux
La restriction de cisaillement
Dans le réseau cellulaire propre à un
tissu ligneux, les mouvements de torsion de chaque cellule
imbriquée sont fortement gênés (blocages par les
cellules immédiatement voisines). Un tel environnement implique donc
la nullité des déformations de cisaillement,
phénomène mécanique souvent appelé "restriction de
cisaillement" (Bodig et al, 1982, Gril, dans Perré,
1998). Dans une telle situation, la double paroi cellulaire
présente alors une rigidité nettement supérieure à
celle d'une double paroi de même structure extraite de son
environnement histologique.
Néanmoins, la prise en compte de cette
restriction de cisaillement dans les modèles multicouches, ne
fait pas l'unanimité. Norimoto et al (1986) considèrent
ainsi que sa prise en compte amène à des prédictions en
désaccord avec leurs résultats expérimentaux.
Description mécanique du renfort cellulosique de
la double cloison par un squelette de microfibrilles
entrecroisées.
La sous couche S2 étant la plus épaisse et la plus
riche en cellulose (exception faite de la sous couche S3, dite couche
gélatineuse, du bois de réaction des feuillus), certaines
modélisations
du comportement mécanique de la double paroi
cellulaire (Guitard et al, 1999, 2002) résument le
comportement mécanique de la cellule à celui d'une unique sous
couche virtuelle dont les caractéristiques ultrastructurales sont
empruntées essentiellement à la sous couche S2.
La cellule ligneuse vue comme une structure cylindrique de
matériau composite
La cellule, réduite à une unique sous
couche S2, (schématisation proposée initialement par Barber
et al, 1964 afin de prédire le retrait longitudinal), est
assimilée à un cylindre à paroi fine constitué
d'un matériau composite fibres cristallines isotropes - matrice
amorphe très spécifique.
La matière constitutive de la cellule virtuelle
est en effet décrite par une matrice amorphe, élastique et
isotrope, de lignine et d'hémicelluloses, renforcée par un
réseau de microfibrilles cristallines isotropes parfaitement
interconnectées (double enroulement croisé, Figure 1.6).
Dans ce type de modélisation (Guitard et al,
1999), initialement proposée pour rendre compte des déformations
de maturation, la composition chimique pariétale de la sous couche S2
n'est pas distinguée selon le type (bois initial, bois final)
de cellule considérée, conformément à certaines
récentes constatations chimiques (Bergander, 2001,
Salmèn communication personnelle, sur Picea abies).
Contrairement aux modèles multicouches
précédents (paragraphe 1.2.7.1), ce modèle à
squelette de fibres, décrivant une double cloison virtuelle, assure
directement la restriction de cisaillement interpariétale par
l'intermédiaire d'un réseau dense de microfibrilles
entrecroisées.
Il n'a en outre pas pour vocation de rendre compte
du comportement mécanique de deux parois mitoyennes (double cloison)
mais plutôt de décrire celui d'un ensemble de cinq cellules
ligneuses par le comportement d'une cellule virtuelle,
énergétiquement équivalente. Ce faisant, il s'agit plus
d'un modèle de tissu cellulaire homogène qu'un modèle de
cellule isolée.
20
Inventaire des facteurs essentiels de la variabilité intra
arbre des propriétés mécaniques des tissus ligneux
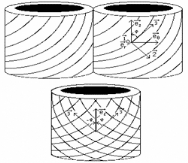
Figure 1.6 Passage de deux cellules adjacentes à
enroulement hélicoïdal de microfibrilles
à une cellule unique virtuelle avec double
enroulement croisé, d'après le modèle de Yamamoto
cité dans Guitard et al, 1999.
Enfin, contrairement aux stratifiés
pariétaux les caractéristiques élastiques de la
double cloison virtuelle sont, par construction, indépendantes de
l'épaisseur du composite i.e. des caractéristiques
géométriques de la cellule ligneuse (Guitard et
al, 1999). Les propriétés élastiques de double
pariétale fournies par ce modèle seront détaillées
dans le chapitre 2.
Le modèle proposé par Guitard et al
(1999) a prouvé sa pertinence quand à la prédiction
des déformations résiduelles de maturation (variations
dimensionnelles de la paroi au cours du processus de maturation
cellulaire) tant longitudinales (DRLM) que tangentielles (DRTM) et
ce à partir des mesures d'angles des
microfibrilles dans S2. Il n'a pas été nécessaire
d'envisager une paroi cellulaire multicouche pour simuler le passage du retrait
au gonflement pour des angles inférieurs à 30 degrés.
Cette modélisation rend ainsi compte de l'inversion de
la DRLM prévue pour un angle situé entre 20 et
30 degrés, et se révèlent apte à la
prédiction simultanée des DRLM et DRTM sur deux essences
résineuses japonaises (Cryptomeria Japonica D. Don, Chamaecyparis
obtusa Endl).
Contrairement à des propositions de
schématisation multicouche de la paroi, il n'a en outre pas
été nécessaire de modifier la constitution chimique
des sous couches cellulaires avec l'angle des microfibrilles pour rendre
compte du comportement de la cellule virtuelle lors du processus de
maturation.
21
Inventaire des facteurs essentiels de la variabilité intra
arbre des propriétés mécaniques des tissus ligneux
| 

