1 ère partie :
A propos du sommeil
5
Nous nous proposons dans cette première partie
d'aborder le sommeil à partir de différentes approches :
- Fonctionnelle, mettant en parallèle l'évolution
des espèces, celle du
sommeil et les grandes fonctions de l'organisme.
- Descriptive centrée sur le sommeil normal de l'homme.
- Neuroanatomiques avec la mise en perspective des structures
impliquées dans la motricité oculaire.
- Expérimentale relative aux interactions entre le sommeil
et la
mémoire
- Théorique à partir de modélisations
neuromimétiques mettant en évidence les aspects thermodynamiques
associés au sommeil, à la mémoire et à
l'adaptation.
- Physiopathologique à partir de l'étude des
conséquences des
privations de sommeil.
- Clinique et séméiologique.
- Standardisée avec la classification des troubles du
sommeil et de la
vigilance.
En dernier lieu nous proposons une synthèse tentant
d'appréhender «la fonction sommeil» en termes
d'homéostasie et d'adaptation.
6
Abord fonctionnel et phylogénèse du sommeil
L'abord fonctionnel du sommeil est un exercice difficile. Le
meilleur moyen de se ridiculiser dit l'un de nos maîtres n'est il pas de
trouver des fonctions au sommeil ? Les définitions fonctionnelles
actuellement admises sont quasiment des lieux communs "Le sommeil est
nécessaire à la prise en compte de l'expérience
vécue et à l'adaptation de la réponse" (Janet - 1889) ,"
Le sommeil a pour objet de permettre la récupération de la
période d 'éveil précédente et de préparer
à la période à venir" (Feinberg - 1974).
Une approche originale, présentée par .M. Krueger
(3) Fig 1, met en perspective l'évolution des organismes vivants,
l'organisation de la réponse de ces organismes aux stimuli
environnementaux et la fonction hypnique. Dans le cas des organismes
unicellulaires il est mis en évidence des modifications fonctionnelles
essentiellement métaboliques (cycles repos/activité) en
réponse aux variations de l'environnement .
Evolution Etat de vigilance Fonction
|
Unicellulaires
|
Stimulus environnemental
|
Métabolique
|
|
Multicellulaires
|
Cycles Repos/Activité Stimulus environnemental
|

|
Cycles Repos/Activité
|
|
Signal humoral
|
Systèmes Ganglionnaires
Stimulus environnemental
Signal humoral
Activité électrique
Systèmes organisés
Stimulus environnemental
Activité électrique
Cycles Repos/Activité
Signal humoral
Cycles Repos/Activité Sommeil
Stimulus environnemental
Organisation Synaptique
Régulation
thermique
Sommeil
Cycles Repos/Activité
Activité électrique Signal humoral
Température
Fig 1 - Cycles repos activités et
phylogénèse du sommeil
Chez les organismes multi-cellulaires on retrouve l'existence de
signal humoral puis électrique en réponse aux fluctuations
environnementales et du cycle Repos/Activité. Le sommeil, en tant que
tel, se différencie de l'état de repos chez les organismes
disposant d'une organisation neurale ganglionnaire. Le sommeil est alors
associé à une fonction d'organisation synaptique. Durant
l'éveil (Repos/Activité) la mise en oeuvre des mécanismes
d'adaptation et de régulation au niveau des réseaux de neurones
se traduit par une perte de la
7
capacité fonctionnelle de réponse à
l'environnement. Le sommeil est alors un moment durant lequel ces
mécanismes d'adaptation peuvent se dérouler de façon
optimale. L'apparition de l'homéothermie (associée au sommeil
paradoxal ) correspond alors à des mécanismes d'adaptation plus
complexes dans lesquels est mise en jeu la thermorégulation.
L'intérêt de cette approche "du bas vers le haut"
est tout d'abord d'envisager le sommeil comme étant une
propriété fondamentale de populations neuronales. Au niveau du
cortex cérébral il est possible de mettre en évidence
l'existence de molécules mises en jeu dans les mécanismes de
régulation du sommeil et augmentées lors des privations de
sommeil (NGF, BDNF, Interleukine-1). A un second niveau, l'hypothèse
d'une fonction cellulaire du sommeil, fait envisager que les fonctions
supérieures du système nerveux central correspondent à un
état évolutif postérieur. Le sommeil ayant alors un
rôle essentiel lié à la régulation de
l'homéostasie par le système nerveux central dans des conditions
particulières (p.ex : réponse aux infections, digestion,
etc...).
Cette approche est aussi celle de McGinty (2) pour qui le sommeil
est un comportement assimilable, sur le plan homéostasique, à
ceux de prise de nourriture ou d'adaptation à l'environnement thermique.
Dans ce schéma, le sommeil est donc dépendant d'un réseau
neuronal localisé dans l'hypothalamus et les structures adjacentes. Une
partie essentielle de ce réseau est composée des neurones
thermo-sensibles. La privation de sommeil a alors pour conséquence
d'induire des perturbations de l'homéostasie et des comportements
associés (chez la souris, perte de poids et des graisses, accroissement
de la prise de nourriture, incapacité à prévenir les
déperditions thermiques). La privation totale de sommeil aboutissant
chez l'animal à la mort en une douzaine de jours dans un tableau de choc
septicémique sans foyer de localisation spécifique.
Fig 2 - Evolution qualitative et quantitative des
états de vigilance en fonction de l'age (d'après
M.J.Challamel)
8
Abord descriptif : le sommeil normal chez l'homme
L'électro-encéphalogramme (EEG) fut la
première mesure utilisée dans l'histoire de la recherche sur le
sommeil. Elle reste la plus importante pour sa description. En plus des
dérivation EEG, l'enregistrement polygraphique classique comporte
l'électro-oculographie (EOG) et l'électro-myographie (EMG) des
muscles du menton.
Cette polygraphie distingue outre l'éveil, cinq stades de
profondeur du sommeil qui font l'objet d'une codification internationale (5).
Les stades sont numérotés de 1, le plus léger, à 4,
le plus profond. Leurs critères de définition sont les suivants
:
- L'éveil : présence d'activité alpha
(8-13c/s) et/ou de fréquences EEG
mixtes de faible voltage.
- Le sommeil lent ou NREM :
Superficiel- stade 1 : fréquences EEG mixtes de
relativement faible voltage avec
prédominance d'ondes thêta
(3,5 à 7,5c/s)
- stade 2 : présence de fuseaux de sommeil (spindles)
à 12-14c/s durant 0,5s
au moins et/ou de complexes K en l'absence
d'une activité lente suffisante pour parler de stade 3.
Profond - stade 3 : tracé EEG occupé par 20%
à 50% d'ondes delta de grande
amplitude à une fréquence
< 2c/s. Un critère d'amplitude est ajouté (=75uV crête
à crête).
- stade 4 : plus de 50% d'onde delta de fréquence =
2c/sec et d'amplitude =
75uV. L'appréciation visuelle a une assez
grande marge d'erreur entre stade 3 et stade 4 ce qui conduit à les
regrouper sous l'appellation de SWS (Slow Wave Sleep) ou de SLP (sommeil lent
profond)
- Le sommeil paradoxal : défini par l'occurrence
simultanée d'un tracé EEG de type 1, de
ou SP, PMO, REM
mouvements oculaires rapides et d'une diminution
générale
complète de l'activité musculaire
tonique de la région mentonnière.
L'organisation de ces états de vigilance n'est toutefois
pas constante lors de la vie de l'individu. L'étude de
l'ontogénèse des états de vigilance met en évidence
à la fois une évolution qualitative, relative à la
répartition de ces états lors du nycthémère et
quantitative quant à leur durée. L'évolution se fait d'une
organisation polyphasique vers une organisation monophasique.
9
De façon plus spécifique la composition d'un
sommeil se décrit à la fois de façon qualitative et
quantitative à partir de l'enregistrement polygraphique.
Les paramètres quantitatifs habituellement retenus sont
:
- La durée cumulée passée en chaque stade
et leur pourcentage par rapport à la durée totale du sommeil.
- Le rapport entre la somme SP (Sommeil Paradoxal) + SLP (Sommeil
Lent Profond : Stade 3 + Stade 4) et le TST (Temps de Sommeil Total)
- Le rapport entre le TST et le temps passé au lit
- Les latences d'apparition du premier Stade 2, du premier Stade
4 et du premier SP
Pour schématiser on peut retenir les pourcentages de
stades suivants :
|
Stade 1
|
Stade 2
|
Stade 3+4
|
Sommeil Paradoxal
|
|
5%
|
50%
|
25%
|
20%
|
Sur le plan qualitatif, l'hypnogramme présente
l'organisation temporelle du sommeil. On peut décrire l'organisation en
4 à 5 cycles d'une période de l'ordre de 90 à 100 minutes
ainsi que l'évolution des différents types de sommeil au fur et
à mesure de l'avancée de la nuit.
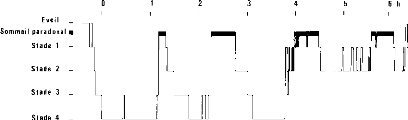
Fig 3 - Représentation des stades de sommeil au
moyen d'un hypnogramme
Il est ainsi possible de repérer, au moyen de
l'hypnogramme, le moment d'apparition de la survenue de différents
phénomènes durant le sommeil (éveils nocturnes,
somnambulisme, somniloquie, énurésie, ...) et de décrire
l'aspect global de celui-ci: fragmenté, instable, avec ou non absence de
certains types de sommeil.
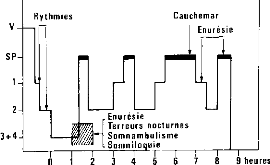
Fig 4 - Moments de survenue des principales
parasomnies
10
Abord neuro-anatomique relatif à l'activité
oculo-motrice
Le sommeil paradoxal est défini par l'occurrence
simultanée d'un tracé EEG de type 1
(fréquences EEG mixtes de relativement faible voltage avec
prédominance d'ondes thêta de 3,5 à 7,5c/s), de
mouvements oculaires rapides et d'une diminution
générale complète de l'activité musculaire tonique
(atonie) des muscles squelettiques.
Le contrôle des muscles de la motricité volontaire
est sous la dépendance des aires corticales motrices qui se
décomposent en aires :
· Pyramidales (aire 4 de Brodmann ou circonvolution
frontale ascendante)
· Extra-pyramidales :
- para-pyramidales de Bucy (aires 6, 4S, 19S et 24S)
- cortico-ponto-cérebelleuses (aires 5, 6, 7, 8, 20)
- cortico-oculo-cephalogyres (aires 8, 9, 19)
- psycho-motrices (aires 6, 8, 9)
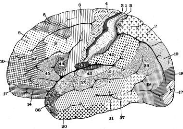
Fig 5 - Vue latérale de l'hémisphère
gauche montrant la cartographie cérébrale de Brodmann
L'influx nerveux élaboré au niveau des aires
corticales est conduit au travers des voies motrices jusqu'aux muscles
effecteurs. Les voies motrices se décomposent globalement en voies :
· Pyramidales, chargées des mouvements volontaires.
Elles mêmes décomposées en faiscean
cortico-médullaire (ou faisceau pyramidal proprement dit) , faisceau
cortico-nucléaire direct (ou faisceau géniculé)
destiné aux noyaux des nerfs crâniens (V, VII, IX, X, XI, XII), en
faisceau cortico-nucléaire aberrant (Déjerine) destiné aux
noyaux de l'oculomotricité (III, IV, VI) et au noyau céphalogyre
du spinal médullaire (XI) innervant le trapèze et le
sterno-cléido-mastoïdien.
· Extra-pyramidales assurant les mouvements
semi-automatiques et associés
· Oculo-motrices, responsables des muscles
extrinsèques et intrinsèques du globe oculaire.
Bien que faisant partie des voies pyramidales et
extra-pyramidales les voies oculo-motrices présentent plusieurs
particularités liées :
· à la spécificité du trajet du
faisceau cortico-nucléaire aberrant qui, à la différence
des faisceaux cortico-médullaire et cortico-nucléaire direct,
n'est pas en relation avec les noyaux du pont (relai des fibres
cortico-ponto-cérébelleuses) mais avec la calotte
pédonculaire et la partie antérieure du faisceau de Reil (pes
lemniscus median).
11
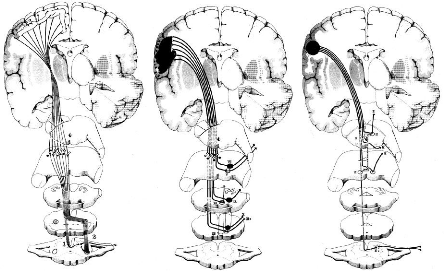
Fig 6 - Faisceaux Cortico-Médullaire,
Cortico-Nucléaires direct et aberrant.
· à la coexistence d'une double musculature oculaire
:
- Extrinsèque responsable des déplacements du
globe oculaire, en accord avec les mouvements de la tête
- Intrinsèque, «réellement»
involontaire, destinée à une meilleure adaptation du cristallin
(l'accommodation) et à une protection de la rétine par le
sphincter iridien (l'irido-motricité).
La motricité extrinsèque constitue le
système oculo-céphalo-gyre qui associe :
· l'oculo-gyrie, assurant les mouvements conjugués
des deux globes oculaires, grâce à
l'action de trois nerfs craniens :
- Le moteur oculaire commun (III)
- Le pathétique ou trochléaire (IV)
- Le moteur oculaire externe ou abducteur (VI)
· la céphalo-gyrie, assurant la rotation de la
tête grâce au nerf spinal médullaire (XI), innervant les
deux muscles trapèze et sterno-cléido-mastoïdien, dont la
contraction unilatérale provoque la rotation de la tête, et dont
la contraction bilatérale entraîne l'extension ou la flexion de la
tête.
Le système oculo-céphalo-gyre utilise deux
neurones :
· Central
- la voie principale volontaire est frontale (aire 8)
- la voie accessoire, semi-volontaire ou réflexe est
occipitale (aire 19)
· Périphérique destiné à
chacun des muscles innervés par les 4 nerfs
oculo-céphalo-moteurs.
La motricité intrinsèque, entièrement
réflexe est sous la dépendance des fibres pupillaires de
12
la rétine, et possède des centres situés
dans les tubercules quadrijumeaux antérieurs. Elle comprend
l'accommodation et l'irido-motricité.
· L'accommodation destinée à mettre au point
le cristallin, comprend 4 neurones :
- rétino-tectal
- tecto-nucléaire
- pré-ganglionnaire
- post-ganglionnaire
· L'irido-motricité déclenchée par
l'excitation lumineuse (réflexe photo-moteur); elle est
aussi le complément de l'accommodation. Celle-ci est sous
la dépendance de deux
systèmes en équilibre :
- irido-constricteur constitué de 4 neurones :
. rétino-tectal
. tecto-nucléaire
. pré-ganglionnaire
. post-ganglionnaire
- irido-dilatateur constitué aussi de 4 neurones mais de
circuit beaucoup plus long car
il met en jeu une voie ortho-sympathique.
. rétino-tectal
. tecto-médullaire
. pré-ganglionnaire
. post-ganglionnaire
1.
Fig 7 - Schématisation des voies oculo-motrices
d'après Bourret et Louis
Muscle dilatateur de l'iris
2. Corps ciliaire
3.
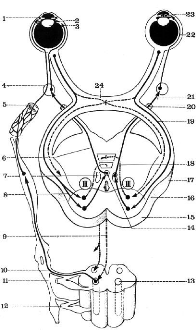
Cristallin
4. Ganglion ophtalmique
5. Artère carotide interne
6. 3ème neurone nucléo-ganglionnaire de
l'accommodation.
7. Noyau d'Edinger-Westphal
8. 1er neurone pupillaire direct de l'accommodation
9. 2ème neurone tecto-spinal de
l'accommodation
10. 3ème neurone spino-sympathique de
l'irrido-accommodation
11. 1ère racine rachidienne dorsale
12. 2ème racine dorsale
13. Centre ciclio-spinal de Budge
14. Noyau de Perlia
15. Tubercule quadrijumeau antérieur
16. Bras conjonctival antérieur
17. Corps genouillé externe
18. Faisceau tecto-spinal
19. 1er Neurone pupillaire direct de
l'irido-constriction
20. Nerf moteur oculaire commun
21. 3ème neurone nucléo-ganglionnaire de
l'irido-constriction
22. 4ème neurone ganglio-musculaire de
l'irido-constriction
23. Muscle constricteur de l'Iris
24. Chiasma optique
III : noyau du nerf moteur oculaire commun
à gauche voies de l'accommodation et de
l'irido-dilatation à droite voies de l'irido-constriction
13
Les fibres issues du noyau spinal médullaire
émergent des cornes antérieures de la moëlle au niveau des 5
à 6 premières vertèbres cervicales. Les racines
pénètrent dans la boite crâniene au niveau du foramen
magnum (trou occipital) et en sortent au niveau du foramen jugulaire (trou
déchiré postérieur) pour former le nerf spinal. Le nerf
spinal (ou nerf accessoire) se divise à la sortie de cet orifice en ses
deux branches terminales :
· La branche interne qui se jette dans le pneumogastrique
à l'extrémité supérieure du ganglion plexiforme.
Elle contribue à l'innervation du voile du palais, du pharynx et du
larynx.
· La branche externe qui innerve le
sterno-cléido-mastoïdien et le trapèze
Sur le plan fonctionnel le nerf spinal assure surtout une
fonction céphalogyre. Toutefois, les muscles innervés agissent
aussi par leur insertion inférieur sur l'orifice supérieur du
thorax. Le nerf spinal entre en jeu dans les mécanismes de l'inspiration
forcée. Le nerf spinal par ses racines bulbaires et avant de se
confondre avec le pneumogastrique prend part à l'innervation du larynx,
c'est à lui que reviendrait essentiellement le rétrecissement de
la glotte et une grande partie de la fonction phonatoire.
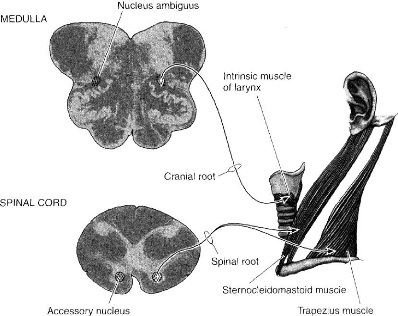
Fig 8 - Branches motrices du nerf spinal (XI)
14
Abord expérimental : sommeil et mémoire
L'apprentissage modifie le sommeil
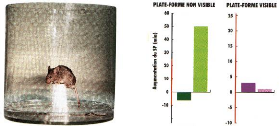
Fig 9 - Test de la piscine
L'effet de l'apprentissage sur le sommeil (6,7) peut être
facilement mis en évidence au moyen de l'expérience de la piscine
: l'animal est mis à l'eau dans une piscine d'eau opaque. Pour reprendre
pied, il doit mémoriser l'emplacement d'une plateforme visible ou non.
Dans le premier cas, il n'y a pas d'augmentation du Sommeil Paradoxal dans les
24h suivant l'entrainement (a droite). Dans le second cas, la durée du
Sommeil Paradoxal augmente de 40% (à gauche).
La privation de sommeil perturbe les apprentissages

Fig 10 - Mémoire et labyrinthe en Y
L'expérience du labyrinthe en Y (8) permet
d'évaluer les conséquences des privations de sommeil sur la
mémoire. La souris étant placée dans un labyrinthe en Y.
On ajoute de la nourriture dans l'une des branches qui lui fait face.
L'animal arrivera t-il à trouver sa nourriture du premier
coup si on le prive de sommeil paradoxal ?
Oui si la branche alimentée est toujours la même (
à gauche, bleu foncé : groupe témoin, bleu clair groupe
privé de sommeil paradoxal).
En revanche, lorsque l'emplacement de la nourriture est
signalé par une lumière et change en essai, le taux de
réponse avoisine les 50%. Ce qui revient à dire que l'animal joue
à pile ou face.
15
Abord théorique : réseaux de neurones et sommeil
Modèle initial de Mac Culloch et Pitts (9 - 1943):
Le neurone est modélisé comme un système
dont la sortie est fonction de la somme pondérée de ses
entrées.
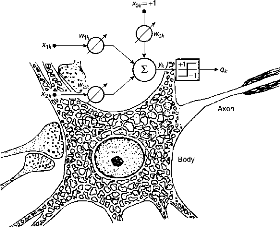
Fig 11 - Modélisation du neurone
Les neurones peuvent être associés entre eux pour
créer des réseaux qui peuvent être
caractérisés par :
· Complexité
· Capacité
· Sensibilité au bruit
· Paradigme fonctionnel
Fig
12
-
|
|
|
|
16
Le perceptron et les réseaux de Hopfield (10)
Il s 'agit d 'un type de réseau caractérisé
par un apprentissage dit supervisé (au moyen d' un module «
enseignant»)
Une fois l'apprentissage réalisé celui-ci
fonctionne sur un mode déterministe
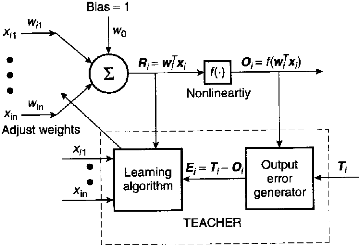
Fig 13 - Réseau de neurones et apprentissage
déterministe
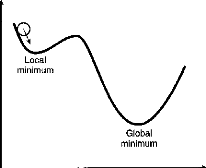
Inconvénient des réseaux déterministes
Le « programme» et la « mémoire» du
réseau sont constitués par les poids synaptiques propres aux
différentes connexions.
Les algorithmes d' apprentissage ont pour effet de minimiser
« l'énergie globale du réseau ». Toutefois, ceux-ci
présentent dans certains cas le défaut de se bloquer sur des
minima locaux de la fonction d'énergie.
Fig 14 - Mémoire et fonction
d'énergie
Le recuit simulé et les réseaux probabilistes
17
Kirkpatrick propose en 1983 (11) d'utiliser la méthode
dite du recuit simulé afin de permettre l'optimisation des
systèmes complexes.
Cette méthode (connue en métallurgie pour la
fabrication des alliages) est constituée de deux phases :
· Une phase de chauffage
· Une phase de refroidissement progressif permettant l
'auto-arrangement du système et la minimisation de son énergie
globale
Recuit et fonction d'énergie

Fig 15 - Fonction d'énergie et recuit
Les machines de Boltzmann (12,13,14)
Il s' agit de modèles fonctionnels de réseaux de
neurones probabilistes proposés initialement par Hinton 1986 et Sussmann
1988.
Ces modèles présentent des capacités d'
auto-apprentissage, l' apprentissage se décomposant en deux phases
alternant successivement.
· Durant la première phase l'ensemble des
associations « entrées sorties » est présenté au
réseau et recuit.
· Durant la seconde phase seules les entrées sont
activées, les sorties pouvant évoluer
librement
Le modèle est-il légitime ?
Il existe chez le mammifère des variations de
température cérébrale associées au sommeil (15 -
Denoyer 1991). De même l'activité neuronale est liée
à la température (Deboer 1995-1998)
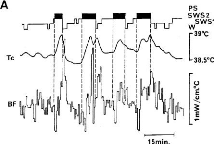
Fig 16 - Evolution de la température corticale
durant le sommeil chez le chat
18
Température et sommeil paradoxal :
On met en évidence, notamment chez les mammifères
l'existence de variations particulières de la température
cérébrale survenant de façon spécifique lors du
sommeil. Ces variations sont essentiellement caractérisées par
une augmentation rapide de la température qui survient de façon
spécifique en début de sommeil paradoxal, puis
décroît progressivement et reste stable jusqu'à la phase
suivante de sommeil paradoxal. L'amplitude des variations observées est
de l'ordre de 0,5°C.
On sait d'autre part que la fréquence de décharge
des neurones est liée à la température, ce qui s'exprime
par le coefficient Q10 (Variation de la fréquence de décharge
pour une augmentation de température de 10°c), dont la valeur est
habituellement de 2 à 3.
La probabilité de décharge d'un neurone est ainsi
d'autant plus élevée que la température est
élevée.
L' effet statistique de la température réelle sur
le fonctionnement neuronal permet d'envisager l'utilisation d'un modèle
formel stochastique. La fonction de recuit dans ce type de modèle permet
de comprendre comment la rétention peut être la conséquence
d'une minimisation des énergies. Cette minimisation de l'énergie
implique un lissage c'est-à-dire, en termes de comportement, un oubli
des états intermédiaires.
Activité EEG à ondes lentes et réseaux de
neurones
Il est possible de proposer un modèle(16) susceptible de
rendre compte du caractère périodique des ondes EEG
observées dans certains états de vigilance (relaxation et
sommeil). Le modèle proposé est constitué d'une
chaîne de neurones électroniques (inspirés du modèle
continu de Hopfield), connectés en cascade. Cette chaîne est
rebouclée sur elle-même.
Chaque neurone, modélisé par un amplificateur de
gain réel positif suffisamment grand associé à une cellule
R-C, apporte un déphasage de: F(ù) = Arctg RC ù.
Dans un tel schéma, le rebouclage du dernier circuit sur
le premier engendre une
oscillation dont la fréquence est:
|
1 2ð
f = tg (
2 ð RC N
|
)
|
|
2 ð 2 ð
( ) ?
Dans la mesure où N est très grand N N
f 1 =
NRC
et
tg
(La fréquence d'oscillation d'un système
composé de N cellules est telle que le déphasage total
apporté par les N neurones doit être de 2 ð. La fonction de
transfert pour chaque cellule est alors telle que : 2 ð / N = Arctg RC
ù)
Il existe alors une relation inversement proportionnelle entre
la fréquence d'oscillation du système et le nombre de neurones
compris dans la boucle. Ceci répond sur le plan descriptif
à la notion classique de synchronisation EEG ou de désactivation,
et sur le plan physiologique suggère qu'il existe une relation entre
fréquences lentes EEG et découplage neuronal
|
|
|
Fig 17 -
Fréquence d'oscillation d'une population
neuronale
19
Bien que très simple, ce modèle nous parait
pouvoir rendre compte : - d'une capacité d'auto-oscillation
(d'auto-oscillation) d'un système de réseaux de
neurones chaînés, - de la cohérence observée de
l'EEG pendant les épisodes Delta du sommeil à ondes lentes. Il
est en accord avec l'hypothèse d'une origine corticale du sommeil
à ondes lentes.
L'évolution de la fréquence de l'EEG lors du
sommeil à ondes (sommeil à ondes)
lentes amène à rechercher un mécanisme de
«couplage sériel» des différentes aires
fonctionnelles.
SOMMEIL A ONDES LENTES ET LOI DE HEBB
La loi de Hebb, relative à l'évolution du poids
synaptique peut être exprimée de la façon suivante (1-2)
:
· Si deux neurones partageant une synapse (pré et
post) sont activés simultanément (de façon synchrone),
alors le poids de cette synapse est augmenté.
· Si deux neurones partageant une synapse sont
activés de façon asynchrone, alors le poids de cette synapse est
diminué.
Durant le sommeil à ondes lentes, l'activité
électrique dans la bande DELTA est caractérisée par une
augmentation de la cohérence intra-hémisphérique en
fonction du stade de sommeil (17). Par ailleurs il a été
proposé l'existence de rapports de phase entre l'activité
fronto-centrale et occipitale (18).
La mise en évidence de rapports de phase et d'une
cohérence élevée de l'EEG durant le sommeil profond
autorise l'application de la 2ème loi de Hebb aux populations neuronales
associées à l'électrogénèse du sommeil
à ondes lentes. On peut alors élaborer l'hypothèse
fonctionnelle qu'une fonction d'oubli (caractérisée par la
diminution des poids synaptiques) serait associée au sommeil à
ondes lentes.
FILTRAGE - RECUIT HYPOTHESE POUR UN PROCESSUS DE
TRAITEMENT DE L'INFORMATION A DEUX ETAPES :
Durant le sommeil à ondes lentes la cohérence
intra-hémispérique de l'activité delta s'accroit avec le
stade de sommeil(17). On met en évidence un rapport de phase entre
l'activité EEG centro-frontale et occipitale (18). Le haut niveau de
cohérence de l'EEG associé à l'existence de ce rapport de
phase permet d'envisager qu'une fonction de filtrage
(caractérisée par la diminution des poids synaptiques) est
associée au sommeil à ondes lentes. Cette fonction agirait comme
un filtre «passe bas» pour les informations mémorisées
drant l'éveil.
Le recuit-simulé (19) est couramment utilisé pour
l'optimisation des apprentissages. Ce processus à deux étapes
optimise l'énergie associée aux patterns. L'existence de
variations de l'activité neuronale en fonction de la températures
permet de proposer qu'une fonction d'optimisation des apprentissages de type
«recuit» est associée au sommeil Paradoxal.
20
Abord physiopathologique des privations de sommeil
Pour Carol Everson (20) les conséquences des privations de
sommeil prolongées chez l'animal sont caractérisées par :
- un hypercatabolisme avec diminution pondérale bien que la prise
alimentaire soit conservée , - un hypométabolisme
cérébral notamment au niveau thalamique, hypothalamique et du
système limbique, - des anomalies endocriniennes avec diminution de la
thyroxine plasmatique secondaire à une perturbation de la
régulation hypothalamo hypophysaire, - une perte de la capacité
de défense immunitaire avec septicémie léthale et
hypothermie. De façon caractéristique, à l'examen
anatomopathologique, on ne retrouve pas de foyer inflammatoire ce qui traduit
une perte des défenses immunitaires. Les germes retrouvés sont
généralement opportunistes.
Chez l'homme, les rares éléments cliniques relatifs
aux privations totales de sommeil nous sont apportés par la connaissance
historique. Dans la Chine ancienne les condamnations à mort s
'effectuaient par privation de sommeil. Lors de l 'exécution des
sentences, les condamnés pouvaient réclamer toutes les
jouissances d 'ici bas, mais, dès que le sommeil les gagnait, leurs
gardiens les maintenaient éveillés par des coups ou autres
sévices. Après une douzaine de jours, la folie survenait, puis,
quatre ou cinq jours après, c 'était la mort dans d 'atroces
souffrances (soit une quinzaine de jours après le début de la
privation totale, le délai est du même ordre que celui
retrouvé chez l'animal).
L'étude des modifications de la réponse immunitaire
secondaire aux privations de sommeil fait actuellement l'objet d'études
avec des protocoles variés (privations transitoires et totales,
privations partielles, etc....).
Lors de privations totales de sommeil sur des durées de
l'ordre de 40 à 64 heures, Dinges (21) retouve une augmentation des
leucocytes portant essentiellement sur les neutrophiles et les monocytes,
l'accroissement du nombre des monocytes se faisant de façon
proportionnelle à la durée de la privation, la normalisation se
faisant avec la récupération, la réponse lymphocytaire
est, quant à elle, difficile à évaluer.
Se basant sur l'hypothèse d'une action immunosuppressive
du sommeil, Pollmächer (22) a étudié l'effet de privations
de sommeil de 40h sur la réponse immunitaire à l'endotoxine. Il
observe seulement une diminution de la réponse fébrile sans
modification de la libération des cytokines ni perturbations
neuro-endocriniennes.
De leur côté, Michael Irwin & Christian Gillin
(23) mettent en évidence une relation entre le sommeil et les aspects
cellulaires de la réponse immunitaire se traduisant notamment par : -
une corrélation entre l'EEG et l'activité des Lymphocytes NK chez
les patients atteints de dépression majeure, - une diminution de la
fonction immunitaire chez les sujets atteints d'insomnie primaire, - une
réduction de l'immunité cellulaire de façon
consécutive aux privations partielles de sommeil, - l'accroissement
nocturne de l'interleukine 6 lié au sommeil, - une diminution de
l'interleukine 6 associée à un accroissement des cytokines
inhibitrices, interleukine-10, chez les patients alcooliques présentant
une perte du sommeil à ondes lentes.
Pour Fehm & al l'interaction sommeil-immunité se
traduit aussi par une perturbation de la réponse immunitaire
observée à la suite de vaccinations. Les sujets mis en privation
de sommeil après vaccination présentent un taux d'Anticorps
circulants moins élevé que ceux qui n'ont pas subi de privation
de sommeil (Fig18).
21
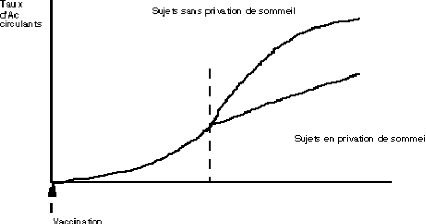
Fig 18 - Effet de la privation de sommeil sur
l'évolution du taux d'Ac circulants (d'après Fehm &
al)
Pour notre part (24) , nous retrouvons, à partir de
l'étude d'une population de 38 000 sujets ayant
bénéficié d'examens de santé, l'existence de
différences significatives des paramètres biologiques
(accroissement des triglycérides) Fig 19. et de la fonction immunitaire
(accroissement des leucocytes) Fig 20. chez les sujets présentant une
plainte relative au sommeil. Les perturbations retrouvées au niveau de
la formule sanguine sont du même ordre que celles observées par
Dinges. L'analyse de la répartition par catégorie
socioprofessionnelle de cette population montre notamment une prévalence
accrue de la plainte concernant le sommeil chez les sujets de sexe masculin en
situation de chômage. Ceci pose de façon aigüe de nombreuses
questions notamment : - sur le rôle des facteurs environnementaux
(stress, temps de transport, chômage ,etc...) susceptibles d'induire une
dyssomnie, - sur les conséquences exprimées en terme de privation
de sommeil des dyssomnies, - sur la prise en charge thérapeutique des
dyssomnies de façon large.
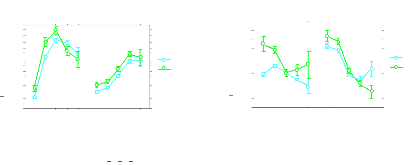
Masculin, inf à 25 Masculin, de 25 à 45
Masculin, de 45 à 60 Masculin, de 60 à 75 Masculin, plus de 75
Féminin, inf à 25 Féminin, de 25 à 45
Féminin, de 45 à 60 Féminin, de 60 à 75
Féminin, plus de 75
Masculin, inf à 25 Masculin, de 25 à 45
Masculin, de 45 à 60 Masculin, de 60 à 75 Masculin, plus de 75
Féminin, inf à 25 Féminin, de 25 à 45
Féminin, de 45 à 60 Féminin, de 60 à 75
Féminin, plus de 75
Graphe des interactions pour Triglycérides
(mmol/l) Effets: TBS * Sexe * Cat Age CES
Barres d'erreurs: #177; 1 Erreurs(s)
standard(s)
Graphe des interactions pour Leucocytes
(giga/l)
Effets: TBS * Sexe * Cat Age CES
Non
Non
Moy. des cellules
Oui
Moy. des cellules
Oui
7,8
7,6
7,4
7,2
7
6,8
6,6
6,4
6,2
6
5,8
1,6
1,5
1,4
1,3
1,2
1,1
1
,9
,8
,7
Fig - 19 Triglycérides et troubles du sommeil Fig 20
-Leucocytémie et troubles du sommeil
De façon plus générale, l'ensemble de ces
interactions sommeil immunité pose de nombreuses questions. Ce d'autant
plus que l'approche "du bas vers le haut " proposée par Krueger met en
évidence l'importance des facteurs environnementaux au sein de cette
régulation complexe. L'état de privation partielle de sommeil est
relativement fréquent au cours de la vie (hospitalisations,
évènements familaux ou professionnels). Il est alors important de
poser la question de l'implication du sommeil dans la morbidité
associée à ces
22
évènements. La mise en place d'une conduite
préventive d'un état de privation de sommeil pouvant alors
être considérée comme un acte d'hygiène (respect des
rythmes de l'individu, limitation des expositions intempestives aux bruits et
à la lumière, prévention de l'anxiété,
.....). En dernier lieu cette approche fonctionnelle éclaire d'un jour
nouveau la théorie de la reprogrammation génétique
développée par M. Jouvet. En effet, autant il est difficile de
concevoir la mise en oeuvre des processus de reprogrammation comme
procédant directement des fonctions hypniques d'apparition relativement
récente dans l'évolution, autant il est facile de les
intégrer comme processus basiques, similaires à ceux de
l'intégration de gènes de resistance par les bactéries
dont l'expression serait très fortement contrôlée par le
paradigme hypnique.
23
Abord clinique des troubles du sommeil :
Celle-ci est articulée en six parties :
- Approche biographique du sujet - Antécédents
médico-chirurgicaux - Habitudes de vie
- Plainte du sujet et anamnèse - Examen clinique
- Examens para-cliniques
Approche biographique du sujet :
Celle-ci s'attache à préciser tout d'abord les
éléments de sa biographie personnelle (date et lieu de naissance,
est-il célibataire, divorcé, quel est son niveau d'études,
travaille-t-il ?, ...) et familiale (profession, antécédents
psychiatriques, toxicomanie, instabilité familiale, ...) des parents,
enfants, frères et soeurs.
Son but est de permettre de déterminer l'environnement
socio-familial du sujet.
Antécédents médico-chirurgicaux :
De façon à permettre de déterminer
l'histoire naturelle de ce qui amène le sujet à consulter. On
s'attachera à rechercher l'existence d'une hypertension
artérielle, de ronchopathie, de troubles digestifs, d'une
obésité, d'une dysthyroïdie, etc... On s'attachera à
préciser l'évolution pondérale, la
régularité des cycles menstruels chez la femme, l'existence de
pollakiurie nocturne.
De même sera recherchée l'existence d'interventions
chirurgicales (même minimes : sutures ) secondaires à un
traumatisme ou à une pathologie médicale (thyroïdectomie,
néphrectomie, etc..).
Sont évoqués ensuite les antécédents
relatifs aux troubles psychiques de l'affectivité, de l'humeur et du
sommeil. Le sujet est il sujet à des crises d'angoisse, à des
épisodes dépressifs ? a t'il utilisé des psychotropes ?
Puis les antécédents relatifs au sommeil:
était-il petit, moyen ou gros dormeur durant l'enfance, l'adolescence,
lors du service militaire pour les garçons ? Etait -il sujet à
des cauchemars, terreurs nocturnes, au somnambulisme durant son enfance ?
Habitudes de vie :
Seront précisées les consommations de substances
à effet psychotrope :
- consommation de café, thé, chocolat (combien,
depuis quand) - consommation de tabac
- consommation de substances non autorisées
- utilisation de tisanes de façon régulière
(passiflore, etc...)
et de façon plus générale la liste de tous
traitements pris de façon régulière
De même les horaires de vie en semaine et lors des
périodes de repos et les aspects subjectifs associés :
- heure du lever (sujet en forme au réveil, temps de mise
en route) - petit déjeuner
- début de l'activité professionnelle («coup
de barre» durant la matinée) - heure du déjeuner
24
- sieste durée, aspect qualitatif, somnolence
post-prandiale
- fin activité professionnelle
- activité au domicile
- heure du dîner
- activité vespérale,
- heure du coucher, endormissement
- éveils nocturnes (nombre et horaire)
- heure du réveil
Ces horaires sont-ils différents en fin de semaine et lors
des périodes de congés ?
Quels sont les horaires spontanés pour lesquels le sujet
se sent bien ?
Plainte du sujet et anamnèse :
Tout d'abord on effectuera le recueil de la plainte
spontanée, de son caractère
handicapant ou non, de l'existence d'une revendication. De
même on s'attachera à savoir si la plainte est motivée par
le sujet ou par l'entourage.
Comment sont apparus les éléments motivant la
plainte? S'agit il d'une évolution progressive ou est il possible de
dater de façon précise l'apparition des troubles ?
Puis dans un second temps on s'attachera à rechercher les
différents symptômes pouvant être associés aux
troubles :
- Y a t'il des difficultés d'endormissement ?
- Le sujet se réveille-t-il la nuit ?
- A t il des comportements particuliers (agitation nocturne,
hypersudation,
parle-t-il)
- Est il réveillé avant de se lever ? a-t-il
tendance à se rendormir sur le petit
matin ?
- Existe-t-il des rêves particuliers à
l'endormissement ou au réveil ?
- Le sujet est il particulièrement sensible aux
émotions ? lui arrive-t'il d'avoir
l'impression d'avoir les «jambes coupées» par
les émotions (joie ou peine)
- Ronfle-t'il ?
- S'endort-il durant la journée ?
- A t-il été victime d'accident ?
Examen clinique
On s'intéresse tout d'abord à la
présentation du sujet, puis à son contact (excitation,
revendication, ralentissement, mimique, etc...)
Un examen général sera ensuite pratiqué,
tension artérielle, allongé puis debout, pulsations,
auscultation, recherche de troubles de la mémoire, des fonctions
supérieures et thymo-affectives. Examen direct ORL.
Examens para-cliniques
Evaluations biologiques en fonction des cas :
- groupage HLA en biologie moléculaire à la
recherche d'un groupe HLA
DR1501/DQ0602 lié à la maladie de
Gélineau
- Dosage de la 6 sulfatoxymélatonine urinaire pour
l'évaluation des troubles de
la synchronisation et des rythmes circadiens.
25
Evaluations électrophysiologiques :
- Polygraphie avec saturométrie ambulatoire pour le
diagnostic des syndromes
d'apnées du sommeil et l'évaluation des rythmes de
sommeil.
- Polysomnographie en laboratoire de sommeil pour
l'évaluation des
hypersomnies.
- Test itératif de latences d'endormissement
- Test itératif de maintien d'éveil
- Actimétrie pour l'analyse des troubles de l'organisation
circadienne
- Enregistrement de la température centrale pour analyse
du rythme de la
température.
- Temps de réaction simple
- Temps de réaction de choix
Evaluations subjectives :
- Agenda de sommeil (recueil sur 21 jours ou plus des horaires de
lever, coucher, temps de sommeil, etc....)
- Auto-Questionnaires :
- Echelles de somnolence d'Epworth
- Echelle de fatigue de Pichot-Brun
- Echelle de matinalité/vespéralité de
Horne
26
Abord standardisé : classification des troubles du
sommeil
L'évocation «troubles du sommeil», amène
fréquemment à l'association d'idées «insomnie».
La classification actuelle se préoccupe davantage des aspects
«fonctionnels» de ces troubles du sommeil et les divise en trois
grandes familles :
- Troubles de l'initiation et du maintien du sommeil
- Les troubles apparaissant à l'occasion du sommeil
- Les troubles liés à l'organisation temporelle du
sommeil
De façon synthétique les troubles du sommeil
peuvent être regroupés selon la classification ICSD (International
Classification of Sleep Disorders) dans le tableau suivant :
|
Troubles de l'initiation et du maintien du sommeil
|
Troubles apparaissant à l'occasion du sommeil
|
Troubles associés à des maladies organiques ou
pshychiatriques
|
|
Intrinsèques
|
Extrinsèques
|
Circadiens
|
Troubles de l'éveil
Troubles de la transition veille sommeil Associés au
sommeil paradoxal
|
Psychoses
Troubles de l'humeur Anxiété
Maladies dégénératives Broncho Pneumopathie
Chronique Obstructive Epilepsie
|
|
Insomnies Hypersomnies S.A.S. M.M.P.I.
|
Grossesse Médicaments Mauvaise hygiène du sommeil
allergie alimentaire
|
Désynchronoses Jet Lag
Travail posté
|
Les classifications telles celles du DSM IV posent
problème essentiellement du fait que les troubles du sommeil sont
étudiés principalement au travers de leur versant comportemental
et non à partir d'un modèle physiologique décrivant les
aspects fonctionnels associés aux états de vigilance. Toutefois
l'approche comportementale à partir des considérations
quantitatives (troubles du sommeil par défaut ou insomnies, troubles par
excès ou hypersomnies) de ces troubles conserve son intérêt
dans la recherche des facteurs étiologiques :
1. Dyssomnies par excès :

L'EXCES DE SOMMEIL
PHYSIOLOGIQUE ANORMAL

INDUIT PATHOLOGIQUE

SECONDAIRE PRIMAIRE PAR TROUBLES
DU RYTHME CIRCADIEN DU SOMMEIL
|
Circadien
|
Insuffisance de sommeil
|
Perturbations
|
Narcolepsie-Cataplexie
|
Syndrôme de retard
|
|
Circasemidien
|
Travail posté
|
respiratoires liées
|
Narcolepsie atypique
|
de phase du sommeil
|
|
Lié à l'âge
|
Vols transmérid iens
|
au sommeil
|
Hypersomnie id iopathique
|
Syndrome d'avance
|
|
de la grossesse
|
Alcool
|
Maladies psychiatriques
|
Impatiences et mouvements
|
de phase du sommeil
|
|
Médicaments
|
Maladies neuro logiques
|
périodiques des membres
|
Syndrôme hypernycthéméral
|
|
|
Maladies infectieuses
|
|
|
|
|
Hypersomnie post-traumatique
|
|
|
27
| 

