TLS, une protéine du spliceosome, est impliquée dans le mécanisme d'action de l'acide rétinoà¯que à travers les régulations post-transcriptionnelle et transcriptionnellepar Eric Le Corvec Université Paris 7 - DEA Biologie des cellules sanguines 2002 |

SOMMAIREb) Préparation de l'ADN plasmidique 11 c) Transfections cellulaires 11 f) Technique de Western Blot 13 g) Technique du retard sur gel 14 1. TLS est un co-activateur du RANC 15 3. Analyse in vivo de l'activité de TLS et de l'acide rétinoïque sur l'épissage alternatif en 5' de E1A 19
I - INTRODUCTIONUn des principaux thèmes d'études de notre laboratoire concerne l'étude de l'hématopoïèse et des leucémies, dans un but de diagnostic, de pronostic et de ciblage thérapeutique. Ainsi, la compréhension des mécanismes moléculaires qui contrôlent la différenciation des cellules myéloïdes est un élément essentiel. L'hématopoïèse est un processus actif, hautement régulé, de prolifération et de différenciation des précurseurs et des progéniteurs hématopoïétiques. Au sein du compartiment actif des cellules souches, un programme de différenciation cellulaire se met en place. Dans un premier temps, la cellule souche perd sa capacité d'auto-renouvellement, puis progressivement sa capacité de prolifération. La cellule engagée dans ce processus va s'orienter vers l'une des voies de différenciation hématopoïétique (granulo-monocytaire, mégacaryocytaire, érythroïde ou lymphocytaire). Les signaux, intra- ou extra-cellulaires, qui mettent en cycle la cellule souche et qui gouvernent secondairement les premières étapes de son déterminisme sont peu connus. Deux modèles sont actuellement proposés pour rendre compte de ce phénomène. Le premier est dit « modèle stochastique » dans lequel ces étapes se font en absence de tout signal extérieur, de façon intrinsèque au programme génétique. Le deuxième modèle est dit « modèle déterministe », où la cellule reçoit de l'extérieur, de façon extrinsèque, un signal qui va déclencher le programme génétique de différenciation. Quel que soit le modèle considéré, ces mécanismes conduisent de façon ultime à l'activation de facteurs de transcription qui activent les gènes responsables de la différenciation cellulaire (Felsenfeld et coll., 1996). C'est vraisemblablement la combinatoire des facteurs de transcription exprimés par la cellule qui va aboutir à l'expression préférentielle des gènes spécifiques de lignée. En d'autres termes, c'est l'expression génique qui influe sur la différenciation. Il a été démontré que les rétinoïdes stimulaient de façon préférentielle la granulopoïèse (Douer et coll., 1982 ; Gratas et coll., 1993). Les rétinoïdes agissent en se fixant sur des récepteurs spécifiques qui sont les récépteurs nucléaires aux rétinoïdes. Les récepteurs nucléaires (Figure 1) appartiennent à une famille de facteurs de transcription qui régulent l'expression des gènes en fonction de la fixation d'un ligand. Les membres de la superfamille des récepteurs nucléaires incluent les récepteurs aux hormones stéroïdiennes, tels que les récepteurs aux glucocorticoïdes (GR) et aux oestrogènes (ER), des récepteurs pour des hormones non stéroïdiennes comme le récepteur aux hormones thyroïdiennes (TR), le récepteur à la Vitamine D3 (VDR), les récepteurs aux rétinoïdes, et également des récepteurs pour divers métabolites lipidique comme les acides gras et les prostaglandines (Green et Chambon, 1986). La superfamille des récepteurs nucléaires comprend également une grande famille de récepteurs dits orphelins lorsque le ligand n'a pas encore été identifié ou n'existe pas. A B C E/F AF-1 AF-2 Activation indépendante du ligand DBD Dimérisation LBD Activation dépendante du ligand Dimérisation D
Figure 1. Structure générale des récepteurs nucléaires Deux classes de récepteurs aux rétinoïdes ont été identifiées : les récepteurs à l'acide rétinoïque (RAR) et les récepteurs au rétinoïde X (RXR). Trois types de RARs (, et ) codés par des gènes distincts localisés sur des chromosomes différents sont présents chez l'Homme, chez la Souris, puis dans d'autres espèces (Giguère et coll., 1987 ; Petkovich et coll., 1987 ; Krust et coll., 1989 ; Zelent et coll., 1989). Les RARs sont capables de fixer deux métabolites actifs de la Vitamine A, l'acide rétinoïque tout-trans (ATRA) et l'acide rétinoïque 9-cis (Allegretto et coll., 1993 ; Allenby et coll., 1993). La comparaison des séquences protéiques des différents RARs et des autres membres de la superfamille des récepteurs nucléaires a permis de les subdiviser en différents domaines possédant des fonctions distinctes. Ces domaines sont plus ou moins conservés d'un récepteur à l'autre et d'une espèce à l'autre (Kastner et coll., 1994). La deuxième classe de récepteurs aux rétinoïdes, les RXRs, lie spécifiquement et uniquement l'acide rétinoïque 9-cis. Les gènes codant pour les trois types de RXRs (, et ) ont été clonés chez la Souris et chez l'Homme. Dans le domaine d'activation des récepteurs nucléaires, on trouve deux séquences portant des fonctions activatrices : AF-1 (domaine A/B), indépendant du ligand et AF-2 (domaine E/F), dépendant du ligand. Ces deux domaines (mais principalement AF-2) peuvent servir au recrutement des co-activateur et des co-répresseurs (Kastner et coll, 1994). La structure des RARs est divisée en six régions fonctionnelles de A à F alors que les RXRs ne possédent pas de région F. Les différents types de RXRs présentent une forte identité de séquence dans leurs régions C et E, mais diffèrent fortement au niveau de leurs régions A/B et D. Cette différence est ainsi spécifique d'un type donné de RXR (Chambon, 1996). Les récepteurs nucléaires exercent leur action par différents mécanismes. Ils peuvent activer ou réprimer des gènes cibles en se fixant directement sur des séquences spécifiques d'ADN appelées éléments de réponse sous la forme d'homo-dimère (exemple des récepteurs stéroïdiens et des RXRs) ou d'hétéro-dimères (exemple des RARs, du VDR et des TRs) avec les RXRs comme partenaires. Ou en fixant d'autres classes de facteurs de transcription (Chambon, 1996). Certains récepteurs nucléaires tels que le TR et le RAR, peuvent réprimer des gènes cibles en absence ou en présence du ligand. Ces effets sont liés à l'interaction des récepteurs nucléaires avec des classes de protéines intermédiaires dont la fonction est soit d'activer (les co-activateurs), soit de réprimer (les co-répresseurs) la transcription. Le complexe transcriptionnel des récepteurs nucléaires à l'AR est dénommé RANC. analyse des régions promotrices des gènes cibles de l'acide rétinoïque a permis de décrire des éléments de réponse, les RARE (Retinoic Acid Response Element). Ces éléments de réponse sont constitués de deux séquences hexa-nucléotidiques conservées PuG(C/T)TCA disposées en répétitions directes (DR), en palindromes ou inversées et séparées par un nombre de nucléotides variant de 1 à 5 (Giguère, 1994). Ainsi, un espacement de 1 nucléotides (DR1) entre les deux motifs consensus AGGTCA définit un élément de réponse de l'homo-dimère RXR-RXR, un espacement de 5 nucléotides (DR5) entre les deux motifs aboutit à un RARE. L'altération de RAR provoque des perturbations au niveau des mécanismes de remodelage de la chromatine. Ces altérations sont dues aux gènes de fusion impliquant le gène RAR lors de translocations chromosomiques responsables de la leucémie aiguë promyélocytaire (LAM3 selon la classification FAB). Il s'agit de la première pathologie maligne humaine répondant à une thérapeutique différenciatrice (pour revue voir Chomienne et coll., 1996 ; Fenaux et coll., 1994). Elle présente ainsi une caractéristique unique par rapport aux autres sous-types de leucémies aiguës (LAM, anomalie de la myélopoïèse). De façon intéressante, il a été révélée que dans 60% des LAM, TLS, une protéine ubiquitaire, était très fortement exprimée (Aman et coll., 1996). D'autre part, des interactions directes entre TLS et le récepteur aux rétinoïdes RXR ont été démontrées (Power et coll., 1998). Ces deux observations nous incitées à étudier les liens éventuels entre TLS et les voies de signalisation aux rétinoïdes dans un contexte hématopoïétique normal et altéré. Le gène TLS fut, en effet, d'abord identifié dans une tumeur maligne à partir de la translocation t(12;16) comme gène codant pour la partie N-terminale de TLS/CHOP, une oncoprotéine de fusion qui est exprimée invariablement associée au liposarcome myxoïde humain (Crozat et coll., 1993 ; Rabbits et coll., 1993). D'autres translocations chromosomiques (à l'origine de sarcomes humains et de leucémies), fusionnent soit TLS soit un gène similaire, EWS à un grand nombre de facteurs de transcription (Zinszner et coll., 1994). Le point commun de ces diverses oncoprotéines de fusion est la présence du domaine N-terminal de TLS ou de EWS. Ce domaine joue un rôle essentiel dans la transformation confirmé par des expériences de transformation utilisant des lignées de souris (Ichikawa et coll., 1999) ou des cellules normales hématopoïétiques (Pereira et coll., 1998). De nombreuses expériences ont permis de mettre en évidence certains rôles de TLS, notamment la réalisation de deux lignées de souris nullizigotes pour TLS. Les souris homozygotes portant une mutation induite de TLS sont stériles avec une importante augmentation d'axes chromosomiques non appariés ou mésappariés dans les spermatocytes préméïotiques. Ces résultats montrent un rôle de TLS dans l'appariement de l'ADN homologue et dans la recombinaison. L'analyse de ces Souris indique que TLS est essentielle pour la survie du nouveau-né, influence le développement lymphocytaire, a un rôle dans la réponse proliférative des lymphocytes B à des stimuli mitogènes, et est nécessaire pour le maintien de la stabilité génomique (Hicks et coll., 2000). La capacité de TLS à se lier à l'ADN est entre autre induite par sa phosphorylation par la PKCII qui est permise par l'activité tyrosine kinase de BCR/ABL. Ces résultats suggèrent que TLS joue un rôle de régulateur dans la leucémogénèse causée par BCR/ABL, favorisant l'indépendance envers les facteurs de croissance et empêchant la différenciation, en modulant l'expression de récepteurs de cytokines (Perroti et coll., 1998) confirmant que sa surexpression dans les LAM puissent être une des causes des désordres observés.
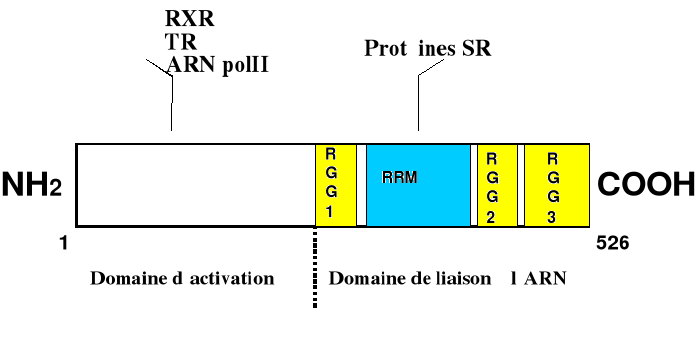
TLS interagit aussi avec l'ARN polII par son domaine N-terminal et est impliquée dans la formation du complexe TFIID. Il pourrait ainsi agir comme un régulateur de la transcription basale participant à la reconnaissance de promoteur transcriptionnel et à l'initiation de la transcription (Yang et coll., 2000). De plus, TLS interagit avec RXR et TR par leur domaine N-terminal (Powers et coll., 1998). Des facteurs d'épissage interagissent aussi avec TLS, l'impliquant dans ce mécanisme. TLS interagit avec Spi-1/PU.1 une protéine ETS capable de réguler la transcription et la maturation ARN dans les cellules myéloïdes (Hallier et coll., 1998). Trois protéines SR, SC35, TASR-1 et TASR-2 interagissent avec TLS par son domaine C-terminal. Figure 2 . TLS est une hnRNP et fait partie de la famille de EWS et hTAFII68.
L'étude de TLS a permis de préciser sa structure (Figure 2). TLS appartient à une famille de protéines incluant EWS (Delattre et coll., 1992) et TAFII68 (Bertolotti et coll., 1996). Le domaine N-terminal des protéines de cette famille est riche en glutamine, sérine et tyrosine, qui sont les acides aminés trouvés dans les domaines d'activation de la transcription. Dans des chimères oncogèniques, l'addition du domaine N-terminal de TLS aux régulateurs transcriptionnels CHOP, FLI-1 ou ERG-1 génère des protéines dont l'activité transcriptionnelle diffère de celle de ces constituants respectifs. De plus, le domaine N-terminal de TLS présent dans ces chimères peut avoir un effet dominant négatif sur la fonction de TLS (provenant de la lignée germinale) comme suggère la récente identification de déterminants oncogéniques dans le domaine N-terminal de TLS. Le domaine C-terminal contient plusieurs motifs : une séquence consensus de ribonucléoprotéine (RRM : ou RNP-CS), des répétitions arginine-glycine-glycine (RGG 2/3), étant la signature des protéines de liaison à l'ARN (Burd et coll., 1994) et un doigt de zinc C2C2. TLS lie l'ARN in vivo ou in vitro (Zinszner et coll., 1997b). In vitro, les ARNs sélectionnés par TLS partagent un motif GGUG. TLS reconnaît un ARN contenant ce motif dans un extrait cellulaire. Chacun des domaines de liaison à l'ARN (les trois boîtes RGG et le motif de reconnaissance de l'ARN) contribuent à la spécificité de l'interaction TLS-ARN (Lerga et coll., 2001). TLS est aussi appelée FUS ou hPOMp75. Cette dernière dénomination a été donnée lorsqu'il fut montré que TLS se lieny à l'ADN simple et double brins. TLS favorise l'hybridation de brins complémentaire d'ADN et l'incorporation d'un oligonucléotide d'ADN simple brin dans une super hélice d'ADN pour former une « D-loop », suggérent que TLS soit impliquée dans la recombinaison homologue (Baetchtold et coll., 1999).
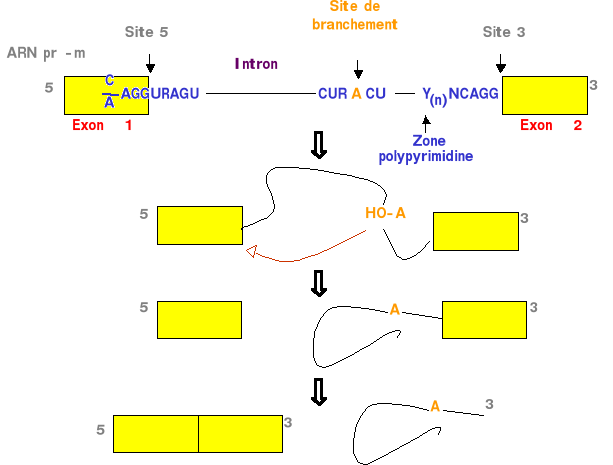
Figure 3. Réaction de transestérification dans le mécanisme de l'épissage de l'ARN pré-m. Chez les Eucaryotes supérieurs, l'expression génique implique la transcription, le « 5' capping », l'épissage de l'ARN pré-m, le « 3' processing », l'exportation de l'ARNm et éventuellement la traduction dans le cytoplasme (Misteli et coll, 2000). L'épissage (dérivation d'un terme de marine qui signifie mettre bout à bout les extrêmités de deux cordages) consiste en une réaction de transestérification éliminant certaines séquences d'un ARN pré-m (les introns) et en laissant d'autres séquences (les exons). Des séquences consensus ont été identifiées aux jonctions exons/introns, elles sont définies comme les sites d'épissage donneur en 5' de l'intron (dinucléotide GU), et accepteur, à l'extrêmité 3' de l'intron (dinucléotide AG) (Breatnach et Chambon, 1981). Les fonctions des snRNA U1, U2, U4, U5, et U6 dans l'épissage sont liées à leur inclusion au sein de complexes composés de snRNA et de nombreuses autres protéines, appelées les snRNP. L'agencement des cinq snRNP avec une cinquantaine de protéines sur la matrice de l'ARN pré-m constitue un complexe ribonucléoprotéique dynamique, le « spliceosome ». Les protéines SR participent à ce complexe, ce sont des facteurs d'épissage essentiels caractérisés par un motif riche en dipeptide sérine-arginine ayant un rôle dans la définition des exons et la sélection des sites alternatifs d'épissage (Valcarcel et coll., 1996). Enfin, la capacité à sélectionner différentes combinaisons d'exons lors de l'épissage, afin de générer des protéines de fonctions distinctes. Ce phénomène d'épissage alternatif est indispensable au développement de nombreux organismes. Environ 60% des gènes seraient exprimés selon un tel mécanisme (Croft et coll., 2000). TLS fait partie de la famille hnRNP. Et elle a été identifiée comme la hnRNP p2 dans un complexe protéique assemblé sur l'ARN pré-m des adénovirus. TLS est engagée dans un complexe avec les hnRNPs A1 et C1/C2 et associée avec plusieurs snRNP du « spliceosome ». TLS est complexée avec des transcrits de l'ARN polII dans des extraits de cellules de HeLa irradiées (Perroti et coll., 1998). En outre, un inhibiteur de l'ARN polII induit la relocalisation de TLS du noyau au cytoplasme, dépéndant de la présence de son domaine C-terminal. Toutes ces proptiétés sont caractéristiques des hnRNPs. TLS fut trouvée associée avec le dinucléotide AG durant l'étape de reconnaisance du site 3' d'épissage et participe à la sélection du site 5' d'épissage alternatif dans l'ARN pré-m du minigène E1A. TLS peut recruter par son domaine C-terminal, deux régulateurs d'épissage de la famille des protéines SR. Ainsi, TLS est un de facteur d'épissage.Lorsqu'elle est surexprimée dans des cellules érythroïdes, TLS induit de préférence l'utilisation du site 5' distal d'épissage, durant la maturation de l'ARN pré-m de E1A (Hallier et coll., 1998). Cette préférence est contrebalancée par Spi-1, suggérant que TLS peut faire partie d'un réseau de protéines impliquées dans la régulation de la maturation ARN. Plusieurs pathologies liées à des altérations de l'épissage ont été caractérisées. Certaines affections comme les amyotrophies spinales héréditaires ou les maladies neurodégénératives liées à la protéine tau, représentent de véritables problèmes de santé publique et donnent lieu à des coopérations fructueuses entre laboratoires hospitaliers et universitaires (Philips et Cooper, 2000). Pour exemple de pathologies impliquant l'épissage alternatif, on retrouve Spi-1/PU.1 dans l'érythroleucémie de Friend et CD44 dans certaines transformations tumorale (Stickeler et coll., 1999). Le rôle de TLS dans la régulation transcriptionnelle n'est pas encore clairement élucidé. TLS augmente la transactivation dirigée par NFB induite par des stimuli physiologiques tels que le TNF, IL-1 et la surexpression d'une kinase induisant NFB. TLS augmente l'activité du promoteur NFB-dépendant d'un gène d'adhésion intercellulaire et du gène de l'IFN. Ces résultats suggèrent que TLS agit comme un co-activateur de NFB (Uranishi et coll., 2001). BUT DU PROJETLe fait que TLS soit surexprimée dans 60% des LAM et qu'il interagisse avec RXR, l'un des principaux partenaires du RANC, nous permet d'émettre l'hypothèse que TLS serait liée à la voie de signalisation de l'AR dans les cellules hématopoïétiques. Nous nous proposons d'étudier deux fonctions de TLS qui pourraient être mises en jeu dans les LAM, en tant que co-activateur de la transcription et son rôle de facteur d'épissage. L'action de TLS (en présence d'AR) comme co-activateur de la transcription est testée dans deux modèles cellulaires, HL-60 et Cos-6, par le dosage de l'expression d'un gène cible sous la dépendance d'un promoteur fixant le RANC. L'appartenance de TLS au RANC est observée par la mise en présence du RANC (RAR, RXR, DR5 avec ou sans AR) et de TLS. L'effet de TLS et de l'AR sur l'épissage sont analysés qualitativement par l'obtention de profil d'épissage alternatif du minigène E1A et l'étude quantitative des isoformes produites. |
|