CHAPITRE III RESULTATS
1-ESSAI PRELIMINAIRES
compte tenu de l'absence totale d'un support bibliographique
traitant la question de la régénération in-vitro
du Scorpiurus ; nous étions amené donc à nous
inspirer , dans une première série d'expérience, que nous
avons qualifiée de préliminaire, d'un certains nombre de travaux
réussis portant sur l'organogenèse et l'embryogenèse
somatique de plusieurs espèces appartenant à la famille des
légumineuses en vue d'établir un protocole expérimental
susceptible de nous conduire à résoudre notre
problématique . Cette problématique qui se résume en fait
à essayer d'obtenir soit des bourgeons néoformés soit des
embryons somatiques et par la même , réussir dans la mesure de
possible, à régénérer des plantes entières
via l'une ou l'autre méthode de micropropagation .
Après l'étude bibliographique , nous nous sommes
parvenues à établir un protocole de travail , qui prenait en
compte les paramètres suivants :
· Un milieu de culture, composé des sels
minéraux et des vitamines de MURASHIGE et SKOOG.
· Des régulateurs de croissance,
représentés essentiellement par le 2,4-D (substance à
effet auxinique) et la BA (cytokinine) employées dans le milieu de
culture, seules ou en combinaison et à différentes doses (0, 0.1
,0.5, 1, 2, 3, 4, 5)
· Des explants de nature et d'origines diverses:
-Des explants de racines, d'hypocotyle et de
cotylédons sont prélevés à la fois à partir
de plantules cultivée in-vitro et dont l'âge varie de 7
à 10 jours et aussi sur de jeunes plantes cultivées
séparément en pots âgées de 30 -35 jours
-Des explants de tige, d'apex, de feuille et de
pétiole sont prélevés de plantes cultivées en pots
âgées de 30 -35 jours .
· Trois génotypes différents appartenant
au genre Scorpiurus ont été choisis de manière
aléatoire. Il s'agit des génotypes V12, B8 et L26.
· En ce qui concerne les conditions de culture, nous
avons retenu à ce que la plupart des expérience seront conduites
à l'obscurité et aussi à la lumière.
1.2 Résultats obtenus
La lecture des résultats des expériences
préliminaires a été faite à trois niveaux de
considération . Le premier, concerne la callogenèse, le second
l'organogenèse et le troisième l'embryogenèse
somatique.
S'agissant de la callogenèse, nous avons
constaté qu'elle s'est déclenchée sur l'ensemble des
explants testés à l'exception des apex qui débourrent et
cela, quelle que soit leur nature ou leur origine . Toutefois, son importance,
semble varier selon l'explant et aussi selon le milieu de culture ( en
particulier sa composition hormonale). En effet, concernant les explants, ce
sont les cotylédons suivies des hypocotyles qui paraissent
répondre le mieux à la callogenèse . Quant aux milieux de
cultures, ils induisent dans leurs majorité, des cals, en dehors de
ceux dépourvus de régulateurs de croissance ou pourvus
uniquement de ( BA) ou de 2,4-D seuls. Les meilleurs réponses sont
obtenues sur les milieux : D3B0,1 ; D0,1 B3 et D1B3.
Les résultats des expériences
préliminaires, nous ont permis de mettre en exergue, l'influence de la
photopériode. L'exposition des cultures sous une photopériode de
16/8 (16 heures de lumière et 8 heures d'obscurité) favorise de
loin la callogenèse que l'obscurité.
Par ailleurs, les plantules ayant servi comme "parc à
bois " ou plantes - mères, apportent aussi leur part d'influence. Ainsi,
nous avons pu relevé que les explants provenant de plantules
cultivées in-vitro réagissent mieux que ceux
prélevés des plantules cultivées en pots .
Ce sont là, les principaux remarques que nous avons pu
relever lors de ces expériences préliminaires .
Au sujet de l'organogenèse, nous avons
remarqué qu'elle s'exprime chez les trois génotypes
testés. Toutefois, comme en callogenèse, son importance est
fonction de la nature l'explant et de la composition hormonale du milieu .
Effectivement, ce sont encore une fois les cotylédons et les hypocotyles
qui expriment des aptitudes organogènes et ce sont aussi les explants
issus de plantes cultivées in-vitro qui réagissent
positivement à ce processus, à l'inverse de ceux
prélevés sur plantules cultivées en pots.
En ce qui concerne les milieux de culture, ce sont ceux
pourvus d'un mélange (2,4-D - BA) qui ont permis l'obtention de
structures organogènes. Ces milieux sont représentés par
le D0,1B3, le D1B3 et le D1B5.
Comme en callogenèse, l'organogenèse est
complètement inhibée à l'obscurité ; un
régime photopériodique de 16/8 ( 16 heures de lumière
et 8 heures d'obscurité ) lui est très favorable (Figure 4).
Concernant l'embryogenèse somatique, les
résultats préliminaires indiquent qu'elle est possible chez le
Scorpiurus. Cependant, elle reste tributaire, comme c'était
le cas en callogenèse et en organogenèse, de plusieurs facteurs
tels que : la nature de l'explant, le génotype et la composition
hormonale du milieu.
Parmi, tous les explants testés, les cotylédons
, les hypocotyles et les racines, sont les seuls à avoir exprimé
des potentialités embryogènes et seulement avec deux
génotypes (le V12 et le B6) sur trois.
Quant à l'influence de la composition hormonale du
milieu, nous avons relevé que, parmi toute la panoplie de milieu de
culture que nous avons testé, seuls les milieux à (2,4-D -
BA), notamment le D3B0,1 et le D0,1B0,5 , possèdent des pouvoirs
inducteurs.
Par ailleurs, il est important de signaler que les
différents types d'explants réagissent mieux à
l'embryogenèse somatique en présence de lumière qu'en
obscurité ( Figure 4).
.
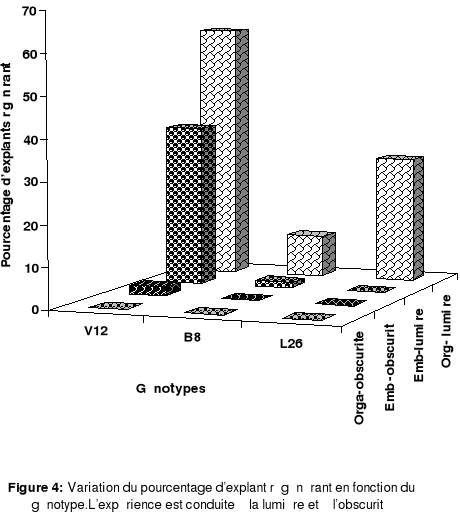
Le temps d'apparition des embryons sur les cals, lorsque
les cultures sont soumises à l'obscurité, prend beaucoup plus
de temps comparativement aux résultats obtenus en présence de la
lumière. Effectivement , les structures embryogènes
n'apparaissent qu'après 60 jours de culture sur le milieu d'induction.
Elles n'atteignent le stade cotylédonnaire qu'au bout du 90
éme jours. Alors qu'à la lumière , elles
apparaissent dés le 15éme jours qui suit
l'ensemencement et atteignent le stade cotylédonnaire au alentour du
25éme jours de culture.
1.3 Coupes histologiques
Nous avons procédé dans
cette partie réalisé des coupes histologiques pour confirmer , si
les structures organogènes ou embryogènes que nous avons
obtenues représentent réellement des bourgeons ou des embryons
somatiques.
1.3.1 Coupes histologiques au niveau des
bourgeons
L'apparition des bourgeons sur les explants hypocotyles,
cotylédons et racines s'est faite de façon progressive.
Néanmoins, il a été possible de distinguer 2 situations.
D'une part, un certain nombre d'explants édifiant des bourgeons
très rapidement (7 à 8 jours) .D'autre part, des
régénérations ne se formant qu'après un temps de
culture plus important (12 à 35 jours ).Il était
intéressant de savoir si le processus d'organogenèse etaient le
même dans les deux cas. Quelques coupes histologiques ont
été effectuées.
· Examen d'explants
régénérant par voie directe (7 à 8
jours)
Les coupes histologiques
permettent d'observer les explants d'hypocotyles ayant conservé une
structure classique. Il semble y avoir une continuité entre le bourgeon
néoformé et les vaisseaux conducteurs, mais il
est impossible d'affirmer qu'elle provient du cambium (Planche 2:A).
· Examen d'explants
régénérant par voie indirecte(12 à 35
jours)
Les explants ont été transformés
par la culture. Les tissus sont plus lâches avec des amas de grandes
cellules très vacuolées. Il y a constitution de cal sur les zone
de blessures et l'intérieur de l'explant. De même,
l'épiderme n'est plus continu mais interrompu, par des cellules de type
cal à cellules de taille variable. On note la présence de
très jeunes méristèmes dont le point
végétatif est identifiable(Planche 2B). Cet ensemble est
très dense fortement colorer. La discontinuité dans les explants
entre les bourgeons néoformés et les tissus conducteurs permet
d'envisager le cal de néoformation à partir de cellules de
cal.
1.3.2 Coupes histologique au niveau des embryons
somatiques
Nous avons regardé la forme et l'organisation
cellulaire des cals embryogènes et des embryons somatiques obtenus,
à travers des coupes histologiques.
Les cellules des cals embryogènes sont souvent
grandes et vacuolisées. On observe quelque fois, la présence
d'amas de cellules différentes du reste du cal. La taille de ces
cellules particulières est inférieure aux autres cellules du cal,
leur contenu est dense et le noyau occupe une grande partie de la cellule.
Elles sont probablement à l'origine des embryons somatique. Les parties
sous-jacentes aux embryons somatiques sont composées de cellules de
grandes tailles(Planche 3:A).
La coupe d'un embryon au stade globulaire ou un pseudo-embryon
sur son cal, nous montre un pédicelle qui le rattache au cal (Planche
3:B). Dans le quel, on trouve pas de liaisons vasculaires qui relieraient le
pseudo-embryon au cal, mais il y à une rupture de l'épiderme
à ce niveau.
1.4 Conclusion
A travers les résultats obtenus lors des essais
préliminaires, nous avons décidé d'orienter notre travail
de recherche sur deux axes essentiels à savoir l'organogenèse
et l'embryogenèse somatique qui paraissent être favorables
à la lumière qu'a l'obscurité. Par ailleurs il
s'avère utile d'utiliser comme source d'explants, ceux issues des
plantules cultivées in-vitro avec les trois génotypes
testées et ayant manifestés des aptitudes embryogénes et
/ou organogénes.
A
B
Planche 2: -Coupes histologiques
longitudinales de bourgeons formé par voie directe après 7
à 8 jours ( A ) et par voie indirecte après 12 à 35 jours
( B) de culture des explants d'hypocotyle du génotype V12 sur milieu de
culture D0,1B3( milieu pourvu de 3mg/l de BA et 0,1mg/l de 2,4-D).
A
B
Planche 3:-Coupes histologiques
longitudinales d'embryons somatiques au stade globulaire (A) et au stade
torpille (B) obtenus à partir d'explant d'hypocotyle du génotype
V 6 sur milieu de culture D3B0,1( milieu pourvu de 3mg/l 2,4-D et 0,1 mg/l de
BA).
ESSAIS D'OPTIMISATION
A la lumière des résultats obtenus au cours des
essais préliminaires , et qui nous ont permis de conclure que la
régénération par organogenèse ou
embryogenèse somatique est possible chez l'espèce
Scorpiurus. Nous avons lancés une série
d'expérience, de grande envergure, dans le but d'apporter des
réponses à un certains nombre de questions que nous nous sommes
posées. Parmi ces questions , on voulait savoir s'il n'existe pas un
problème de répétabilité ou de
reproductibilité des expériences qui peut poser dans notre cas.
On voulait aussi déterminer les possibilités d'optimisation de la
production de bourgeons ou d'embryons somatiques et en dernier, voir
l'éventualité de régénérer des plantes
entières via l'organogenèse ou l'embryogenèse, autrement
dit, tester la convertibilité des bourgeons ou embryons somatique en
plantes.
Pour cela, nous avons intégré, dans ces
expériences, d'une part des régulateurs de croissance autres que
ceux utilisés lors des essais préliminaires. Il s'agit de l'AIA
et de l'ANA pour les auxines et de la Kinétine pour les cytokinine et
d'autre part de nouveaux génotypes (V13, V6 ,B6,B1 et L3) dans
l'intention d'élargir, le plus possible, la variabilité
génétique de l'espèce étudiée et augmenter,
ainsi, les chances de tomber sur un génotype à fort potentiel
organogène et ou embryogène.
2.1 Aptitude à la callogenèse
Lors de cet essai , nous avons testé l'effet que peut
avoir l'apport seul puis combiné des auxines ou des cytokinines, sur
l'aptitude à la callogenèse chez les huit génotypes de
Scorpiurus choisis, en se servant des explants ayant montré des
capacités organogènes et embryogènes lors des essais
préliminaires. Les cultures sont conduites sur milieu de base MS
contenant différentes combinaisons hormonales.
Le suivi de la callogenèse porte à la fois sur
la détermination du pourcentage d'explants callogènes,
l'estimation quantitative des cals formés, et sur la description des
caractéristiques morphologiques des cals (texture et couleur) .
2.1.1 Influence des régulateurs de
croissance
Les régulateurs de croissance testés
sont les auxines, représentées par le 2,4-D, l'AIA et l'ANA
et les cytokinines , représentées par la BA ou BAP et la
Kinétine. Ils sont apportés seuls ou combinés entre eux ,
à différentes doses (0,1; 0,5 ; 1; .. . . . . .15; 20; 25 ;30 ;
35 ; 40).
Action des auxines ou des cytokinines
seules
De manière générale, l'usage des
auxines seules, à faibles concentrations dans le milieu d'induction
s'est révélé inefficace à la callogenèse
quelque soit le génotype ou le type d'explant utilisé .
Cependant, leur apport à forte concentrations semble déclencher
une légère callogenèse , souvent localisée au
niveau des zones de blessures des explants. C'est le cas du 2,4-D ou de l'ANA
et l'AIA lorsqu'ils sont fournis, au milieu, respectivement aux doses de (5
mg/l à 10 mg/l) et (10 mg/l à 40 mg/l). L'emploi de
concentrations plus fortes devient létale.
Quant à l'apport des cytokinines seules, dans le milieu
d'induction, quelle que soit la dose, la nature de l'explant , ou le
génotype utilisé, il se montre sans effet aucun sur la
callogenèse.
Action de la combinaison auxine-
cytokinine
Les combinaisons hormonales testées, lors de ce
travail, sont du type (2,4-D x BA; 2,4-D x Kin ; AIA x BA ; AIA x Kin ;
ANA x BA et ANA x Kin).
Les résultats du tableau 3, représentant
l'influence de la nature et de la concentration des régulateurs de
croissance sur l'aptitude à la callogenèse des cotylédons
de plusieurs génotypes, montrent clairement que l'apport combiné
des auxines et des cytokinines ne peut être que bénéfique
et par conséquent très stimulant à la callogenèse.
En effet, la callogenèse a été induite, mais à des
degrés différents, sur l'ensemble des milieux testés et ce
sont les combinaisons à base de 2,4-D qui semblent développer les
meilleurs pouvoir inducteur. La callogenèse est, davantage,
améliorée lorsque l'association ou la combinaison regroupe le
2,4-D et la BA.
On note par ailleurs que le pourcentage callogène des
explants devient plus important, lorsque le rapport auxine/cytokinine , dans
le milieu, est supérieur à 1.
On remarque aussi que la présence de fortes doses de
kinétine, dans le milieu, permet souvent l'obtention de cal à
texture friable et à aspect chlorophylliens (coloration vert
pâle)( Planche 4:A). La présence de BA,
quant à elle , semble favoriser la production de cals
à texture compacte et à coloration verdâtre (Planche 4:B),
bien qu'il y ait parfois présence de tâches rougeâtres .
Tableau 3 : Variation de l'aptitude
à la callogénèse en fonction de la composition hormonale
du ( quelque soit le génotype testé). Les résultats sont
obtenus après 30 jours de culture des explants cotylédons .
Taille des cals : (+) : (0 - 0.5)cm ; (++) :
(0.5 - 1)cm ; (+++) : (1 - 1,5) cm.
|
Composition hormonale du milieu
|
Pourcentage d'explants callogènes
|
Couleur des cals
|
Texture des cals
|
Taille des cals
|
|
N4B0,1
N4K1
D3B0,1
A20K11
A20B1
D2K1
|
43.57
30.62
85.75
42.50
40.87
60.62
|
Vert
Vert
Vert
Vert
Vert
Vert
|
Compacte
Compacte
Compacte
Compacte
Compacte
Compacte
|
+
+
+++
++
++
+++
|
|
N1B3
N0.1K11
D0.1B3
A1B3
A4K11
D0.1K1
|
30.00
20.87
91.75
19.00
12.37
36.37
|
Vert
Vert pâle
Vert
Vert
Vert pâle
Vert
|
Compacte
Friable
Compacte
Compacte
Friable
Compacte
|
++
++
+++
++
++
++
|
2.2.2 -Influence de l'explant
Les résultats de nos expériences, nous ont
révélé que la callogenèse dépendait aussi de
la nature des explants. Dans notre cas, ce sont les explants de
cotylédons suivis des hypocotyles qui réagissent le mieux
à la callogenèse. Leur pourcentage d'explant callogène
est presque similaire et il est de l'ordre de 45% à 100%.. Les racines,
quant à elles, paraissent développer de faibles capacités
callogène avec un pourcentage oscillant généralement
entre 13 % et 40%.
La callogenèse se déclenche après les
cinq premiers jours de culture qui suivent l'ensemencement. Les explants
commencent d'abord par connaître un léger gonflement,
accompagné de brunissement, au niveau des zones d'excisions. Les cals
font, souvent, leur apparition au niveau de ces zones et sur les facettes, des
explants, se trouvant en contact avec le milieu et se généralise,
par la suite, pour couvrir la totalité de l'explant. Le
développement de la callogenèse devient plus important au
delà du 10ème ou du 12 ème jours de culture
On note par ailleurs, que la croissance des cals varie aussi
d'un type d'explant à un autre. Ainsi, nous avons pu constaté que
les cals provenant des explants de cotylédons, malgré leur
apparition tardive, connaissent une croissance plus rapide et donc plus
importante que ceux produits par le reste des explants : hypocotyles et
racines.
2.2.3-Influence du génotype.
Les résultats préliminaires ne nous ont pas
permis d'avoir une idée précise sur l'effet génotypique et
ses conséquences sur la callogenèse, compte tenu du nombre
très restreint de génotypes soumis à l'expérience.
L'élargissement des expériences à d'autres
génotypes est devenu donc nécessaire. Ainsi, huit
génotypes de Scorpiurus ont été choisis, afin de
tester leurs aptitudes à la callogenèse, en se servant d'un
milieu (D0,1B3) ayant montré de bon pouvoir inducteur, lors des
résultats préliminaires (l'annexe 3).
Les résultats du tableau 4, présentant les
aptitudes à la callogenèse, montrent que, pour un explant
donné, l'ensemble des génotypes testés développent
une callogenèse. Toutefois son, importance change d'un génotype
à l'autre. Les génotypes de l'espèce Scorpiurus
vermiculatus ( V12, V13 et V6) et Scorpiurus muricatus ssp
sulcatus donnent les meilleurs pourcentages callogènes.
Tableau 4: Variation de l'aptitude à
la callogénèse de huit génotypes de Scorpiurus
sur milieu D0,1B3. Les résultats sont obtenus après 30 jours de
culture des explants cotylédons .
Taille des cals : (++) : (0.5 - 1)cm ; (+++) :
(1 - 1,5) cm.
|
Génotypes
|
Pourcentage d'explants callogènes
|
Couleur des cals
|
Texture des cals
|
Taille des cals
|
|
V12
V6
V13
B1
B8
B6
L26
L3
|
100.00
98.00
96.00
76.00
83.00
85.00
96.00
100.00
|
Vert
Vert
Vert
Vert
Vert
Vert
Vert
Vert
|
Compacte
Compacte
Compacte
Compacte
Compacte
Compacte
Compacte
Compacte
|
+++
+++
+++
++
++
++
+++
+++
|
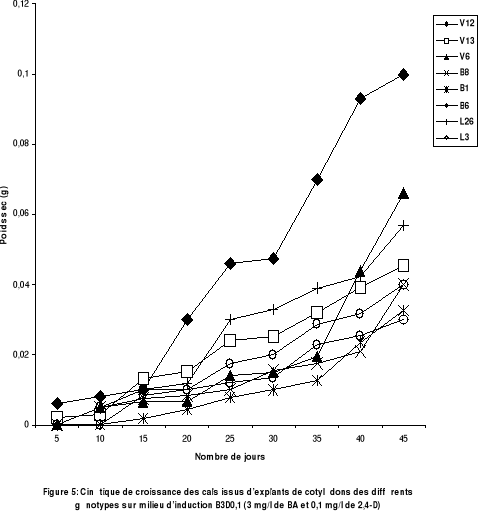
Le suivi de la callogenèse, en établissant une
cinétique de croissance des cals obtenus, avec les explants de
cotylédons des huit génotypes, sur milieu d'induction D0,1B3 (
fig 5), reflète bien l'existence d'une variabilité
génotypique. En effet, avec les génotypes de l'espèce
Scorpiurus vermiculatus, en particulier le V12, la croissance des cals
est plus importante, comparativement aux génotypes de l'espèce
Scorpiurus muricatus..
2.2.4-Conclusion
L'ensemble des génotypes et explants testés ont
manifesté des aptitudes callogènes. Le pourcentage, la texture,
la couleur et la taille des cals dépend de la composition hormonale du
milieu, de l'explant et du génotype. En effet , l'usage des auxines ou
des cytokinines seules, dans le milieu d'induction, ne favorise pas la
callogenèse et les meilleurs résultats sont obtenus avec les
explants cotylédons et hypocotyles des génotypes de
l'espèce Scorpiurus vermiculatus et Scorpiurus muricatus
ssp sulcatus sur le milieu de culture D0,1B3.
A
B
Planche 4: Différents types de cals
retenus après 20 jours de culture des explants de racine sur milieux
d'induction (MS) contenant 4 mg/l d'AIA et 11mg/l de Kinétine (A);
3mg/l de BA et 0,1mg/l de 2,4-D(B)
| 

