Evaluation des effets de la toxicité du cuivre et du cobalt sur la croissance du blé tendre (triticum aestivum) dans la région de Lubumbashi (RD Congo)par Jonas NTAMBWE Université de Annaba - Diplôme de Master 2023 |

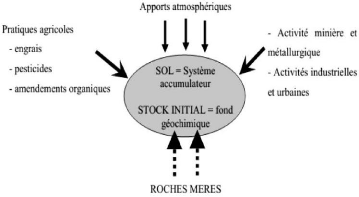
IntroductionLes éléments traces métalliques sont naturellement présents dans les sols à des concentrations faibles inferieures à 0,1% de la composition de l'écorce terrestre (Baize, 1997 ; Kabata-pendias et al., 2001). Bien que certains de ces éléments traces métalliques soient essentiels pour les plantes, ils deviennent tous toxiques lorsqu'ils sont présents à des fortes concentrations (Kupper et al., 2000 ; Baize et al., 2001). La contamination des sols par les éléments traces métalliques peut avoir deux conséquences majeures. D'une part, elle peut favoriser la dispersion des éléments traces métalliques dans l'environnement vers les eaux de surface par érosion, vers les eaux souterraines et les nappes par lixiviation-lessivage et, enfin, par transferts dans la chaine alimentaire par l'intermédiaire des animaux et plus encore des végétaux (Adriano, 2001). D'autre part, la contamination des sols par les éléments traces métalliques peut affecter les différentes fonctions du sol, celles contribuant aux services écosystémiques, et en particulier sa fonction de production en zone agricole (Adriano, 2001). En République Démocratique du Congo, les concentrations de Co et de Cu dans certains régions contaminées peuvent atteindre des milliers de fois supérieures à celle du sol normal (Lange et al., 2016). Le cobalt (Co), qui n'a pas été reconnu comme un élément nécessaire aux plantes bien qu'il ait une fonction biologique ou physiologique distincte dans les légumineuses, constitue une menace sérieuse pour la croissance et le développement des plantes une fois qu'il est disponible en grande quantité dans les sols (Chatterjee, 2003 ; Gopal et al., 2003 ; Karuppanapandian et Kim, 2013 ; Lwalaba et al., 2017a ;b). D'autre part le cuivre (Cu) est l'un des micronutriments requis en très petites quantités par les plantes, une quantité excessive de Cu dans le substrat peut limiter la croissance des racines en brulant leurs extrémités, causant ainsi une croissance racinaire latérale excessive (Bloodnick, 2014). Nombreuses espèces végétales peuvent être atteintes par la pollution de métaux lourds, notamment les espèces céréalières. Aniol et Gustafon (1984) ont montré que les différentes céréales ne réagissent pas de la même façon à la présence des métaux lourds. Ces deux auteurs ont classés, en 1984, le seigle comme la céréale la plus tolérante, le blé présentant une tolérance 2 intermédiaire et l'orge comme la plus sensible. Le blé (Triticum aestivum) a été choisi durant cette étude comme céréale. Cette étude a pour objectif principal d'évaluer les effets de la toxicité du cuivre et du cobalt sur la croissance du blé tendre dans la région de Lubumbashi. Spécifiquement l'étude présente a pour objectif de : ? Evaluer la toxicité des différentes doses croissantes du Cu sur la croissance du blé tendre ; avec comme hypothèse la croissance du blé tendre serait fonction des différentes concentrations du Cu ? Evaluer la toxicité des différentes doses croissantes du Co sur la croissance du blé tendre ; avec comme hypothèse la croissance du blé tendre serait fonction des différentes concentrations du Co La présente étude est subdivisée en quatre chapitres outre l'introduction et la conclusion notamment : le chapitre 1 consacré à la revue de la littérature ; le chapitre 2 présente le milieu, matériels et méthodes, le chapitre 3 présentation des résultats obtenues et le chapitre 4 discussion des résultats. (a) Origine naturelle Les éléments traces métalliques sont présents naturellement dans les roches, ils sont libérés lors de l'altération de celles-ci pour constituer le fond géochimique 3 Chapitre 1. Revue de la littérature 1.1. Eléments traces métalliques 1.1.1. Définition des « éléments traces métalliques » D'un point de vue purement chimique, les éléments de la classification périodique formant des cations en solution sont des métaux. D'un point de vue physique, le terme « les ETM » désigne les éléments métalliques naturels, métaux ou dans certains cas métalloïdes (environ 65 éléments), caractérisés par une forte masse volumique supérieure à 5 g.cm3 (Adriano, 2001) (Tableau 1). D'un autre point de vue biologique, on en distingue deux types en fonction de leurs effets physiologiques et toxiques : métaux essentiels et métaux toxiques.
Les éléments traces métalliques, implique aussi une notion de toxicité. Le terme « éléments traces métalliques » est aussi utilisé pour décrire ces mêmes éléments, car ils se retrouvent souvent en très faible quantité dans l'environnement (Baker et Walker, 1989). 1.1.2. Origine de la contamination des sols par les métaux lourds Le problème principal avec les métaux lourds comme le plomb, le cadmium, le cuivre et le mercure est qu'ils ne peuvent pas être biodégradés, et donc persistent pendant de longues périodes dans des sols. Leur présence dans les sols peut être naturelle ou anthropogénique. 4 (Bourrelier et Berthelin, 1998). La concentration naturelle de ces métaux lourds dans les sols vaire selon la nature de la roche, sa localisation et son âge. (b) Origine anthropique Cependant, la source majeure de contamination est d'origine anthropique. Au cours des décennies dernières, l'apport des éléments traces métalliques au sol dans le monde s'est étendu ; à l'heure actuelle on l'estime à 22000 tonnes de cadmium, 939000 t de cuivre, 783000 t de plomb, et 1350000 t de zinc (Singh et al., 2003). Les principaux types de pollutions anthropiques responsables de l'augmentation des flux de métaux, sont la pollution atmosphérique (rejets urbains et industriels), la pollution liée aux activités agricoles et la pollution industrielle.
Figure 1. Origine des éléments traces métalliques dans le sol d'après Robert et Jules 1999 La pollution atmosphérique résulte des activités industrielles (rejets d'usine) et urbaines (gaz d'échappement, etc....). Il faut distinguer les apports diffus aériens d'origine lointaine des apports massifs localisés d'origine proche. Dans les apports diffus sont classés les poussières et aérosols provenant des chauffages ainsi que des moteurs d'automobiles. Les apports massifs localisés résultent d'apports anthropiques accidentels liés aux activités industrielles sans protection efficace contre la dispersion dans l'environnement (Baize, 1997). Certaines pratiques agricoles sont à l'origine de l'introduction de métaux lourds dans le sol. Les produits destinés à améliorer les propriétés physico-chimiques du sol sont souvent plus riches en métaux lourds que le sol lui-même par exemple les engrais, les composts et les boues de station d'épuration (Robert et Juste, 1999). La pollution industrielle provenant des usines de production de l'activité humaine tels que les matières organiques et graisses (industries agro-alimentaires), les 5 produits chimiques divers (industries chimiques), les matières radioactives (centrales nucléaires, traitement des déchets radioactifs) et la métallurgie (Godin et al., 1985). Les déchets miniers et les terrils industriels sont une source particulièrement importante de pollution par le zinc, le plomb et le cadmium. Le rôle des pratiques industrielles et agricoles dans la contamination des sols doit être pris en compte : cela concerne une grande partie du territoire. Leur accumulation et leur transfert constituent donc un risque pour la santé humaine via la contamination de la chaîne alimentaire, mais aussi pour le milieu naturel dans son ensemble (Bourrelier et Berthelin, 1998). 1.1.3. Mobilité et biodisponibilité des éléments traces métalliques La toxicité d'un métal dépend de sa spéciation (forme chimique) autant que des facteurs environnementaux (Babich, 1980). Dans le sol, les éléments traces métalliques peuvent exister sous forme d'ion libre ou sous forme liée à des particules de sol. Cependant, un métal n'est toxique pour les organismes vivants que s'il est sous forme libre ; il est alors biodisponible. Comme tout élément chargé positivement, les cations métalliques peuvent interagir dans le sol avec toute particule organique ou minérale chargée négativement. De l'équilibre entre les formes libres et fixées de l'ion va dépendre sa biodisponibilité, directement liée à sa toxicité. Enfin, la biodisponibilité des métaux lourds varie en fonction de plusieurs facteurs du sol. Parmi lesquels, la capacité d'échange de cation (CEC), le pH, le potentiel redox (Eh), la teneur en phosphate disponible, la teneur en matière organique et les activités biologiques. Tableau 1. Mobilité et biodisponibilité
du Cu et Co dans les écosystèmes
métallifères
,, 6 1.2. Le Cuivre 1.2.1. Le Cuivre dans l'environnement Un des premiers métaux utilisés par l'homme est le cuivre du fait qu'on le trouve dans des gisements minéraux de beaucoup de régions du monde, et bien qu'il puisse être facilement travaillé, il a une résistance élevée à la corrosion (Angeles Cid, 1995). Le Cuivre est surtout présent dans la croute terrestre sous forme de sulfure de Cuivre (La chalcocite, Cu2S), de sulfure de cuivre-fer (la chalcopyrite, CuFeS2), de bonite (CuFeS4), et aussi comme des minéraux oxydés tels que les carbonates, sulfates et le cuivre des hydroxyles-silicates (Kiaune et Singhasemanon, 2011). Le Cuivre se retrouve dans l'eau de surface, l'eau souterraine, l'eau potable et l'eau de mer mais aussi il est initialement présent sous forme de complexes ou matière particulaire (A TSDR, 2002). 1.2.2. Toxicité du Cuivre Le cuivre est un oligo-élément essentiel pour le fonctionnement des plantes, des microorganismes et des animaux en raison de sin exigence pour de nombreux processus métaboliques particuliers (ICA, 1995). En particulier, il est essentiel pour de nombreuses fonctions biochimiques, comme par exemple la catalyse de réactions d'oxydo-réduction dans le cytoplasme, la mitochondrie et le chloroplaste des cellules (Fargasoya, 2004), ou bien comme un transporteur d'électrons durant la respiration (Yruela, 2009). Le cuivre participe aussi dans la lutte des espèces réactives de l'oxygène comme la Cu/Zn superoxyde dismutase (SOD) qui convertit l'anion superoxyde en dioxygène et peroxyde d'hydrogène (ICA, 1995). Dans le sol, le Cu2+ existe sous plusieurs espèces chimiques, mais il est principalement absorbé comme un élément nutritif par les plantes sous forme de Cu2+ (Maksymiec, 1997). L'accumulation du Cu2+ peut atteindre des concentrations de 20 et 30 ug.g- 1 de poids sec (Pham et al., 2013). La caractérisation primaire de toxicité du Cu2+ est l'inhibition de l'élongation des racines et la croissance de la plante (Tsay et al., 1995). Les effets du Cu2+ provoquent d'autres symptômes comme la nécrose, la chlorose et la décoloration des feuilles (Yruela, 2009). Le changement redox du Cu+ en Cu2+ induit la formation d'espèces oxydées radiculaires comme l'oxygène singulet (02-) et le radical hydroxyle (OH), et ces espèces oxydatives provoquent des dommages aux macromolécules biologiques, comme l'ADN et l'ARN, les protéines, les lipides et les glucides (Lombardi et 7 Sebastiani, 2005 ; Yurekli et Porgali, 2006). Il a aussi été démontré que la toxicité du Cu2+ induit une réduction de l'activité photosynthétique par une baisse du rendement quantique du photosystème II (Alaoui-Sosse et al., 2004 ; Pesko et Kralova, 2013). Egalement, les êtres humains sont exposés au cuivre par inhalation de particules de Cu2+, l'ingestion d'eau ou de nourritures contaminées par le cuivre. Cependant, la toxicité du cuivre pour l'être humain est relativement faible par rapport aux autres métaux tels que le Cadmium, le mercure, le plomb et le chrome. Les effets toxicologiques qui peuvent être induites par les ions cuivre (Cu2+) peuvent provoquer des dépôts dans le cerveau, le foie, la peau, le pancréas, et le myocarde (Davis et al., 2000). 1.3. Le Cobalt 1.3.1. Mobilité et biodisponibilité en milieu terrestre Sols Le cobalt stable est présent dans les sols à une concentration variant de 1 à 50 mg.kg-1 sec (Hamilton, 1994) ; des concentrations élevées ont été observées localement sur des zones hyperminéralisées (jusqu'à 5 000 mg.kg-1 sec) ou sur des sols formés sur serpentine (Coughtrey et Thorne, 1983). En France, les concentrations observées vont de 2 à 23 mg.kg -1 sec avec des valeurs extrêmes allant jusqu'à 150 mg.kg- 1 sec dans des zones de « métallotectes » à fortes minéralisations, notamment dans le Morvan (Baize, 2000). Les sols agricoles de pâture sont souvent déficients en cobalt (< 1 mg.kg-1 sec) et doivent alors être supplémentés en cet oligoélément pour assurer une nutrition correcte du bétail qui s'y nourrit (Hamilton, 1994). De nombreuses études de spéciation et de sorption-désorption du cobalt ont été effectuées sur des milieux simples (oxydes métalliques en particulier) et pour des conditions applicables à certains stockages profonds (Brooks et al., 1998 ; Brooks et Carroll, 2003 ; Khan et al., 1995 ; Wang et Papenguth, 2001 ; Wang et al., 2003). En revanche, les études concernant le comportement du cobalt dans des sols restent plus globales. En termes de spéciation, dans les conditions de pH et de potentiel redox similaires à celles des sols « moyens », le cobalt serait essentiellement sous forme de Co2+. Au-delà de pH 9,5, les hydroxydes de cobalt seraient dominants et tendraient à précipiter ; les sulfures n'apparaîtraient qu'en conditions réductrices (Eh < - 300 mV à pH 7) (Krupka et Serne, 2002 ; Deltombe et Pourbaix, 1966). Le cobalt forme des complexes très stables avec les ligands organiques, notamment les acides humiques et fulviques (Glaus et al., 2000), ce qui augmenterait sa mobilité. Il est reconnu que la chimie de l'élément est étroitement liée à celle du fer et du manganèse et qu'il est étroitement associé aux oxy-hydroxydes 8 métalliques. Sa forte affinité pour les oxydes de fer induit une augmentation de sa mobilité en milieu réducteur, notamment hydromorphe ; cependant, le contrôle de sa mobilité dépend des oxy-hydroxydes métalliques, de la concentration en Ca2+, mais aussi de la présence de ligands organiques (Bresson et al., 2009 ; Krupka et Serne, 2002). La sorption du cobalt sur des phases minérales pures est modérée et décroît avec le pH en l'absence de ligands organiques, alors qu'en présence de complexant organiques la sorption du cobalt diminue par formation de complexes anioniques (Krupka et Serne, 2002 ; Wang et al., 2002). Bien que dans les sols, le cobalt soit moyennement mobile, ses formes complexées sont très mobiles et facilement disponibles pour les plantes. Il serait majoritairement, soit faiblement adsorbé sur les argiles, en liaison avec les oxydes de fer et de manganèse, soit complexé par les acides humiques. Seuls 5 à 10 % du cobalt total d'un sol sont hydrosolubles ou facilement extractibles (Colle et al., 1991 ; 1996). Une évolution notable des fractions échangeables et des fractions très fortement fixées a été montrée par des extractions séquentielles sur des sols de rizière : le cobalt de ces deux fractions tendrait à diminuer en quelques mois, au profit d'une augmentation du cobalt associé aux oxydes de fer (Tagami et Uchida,1998). Par ailleurs, l'augmentation de la teneur en argiles s'accompagne d'une augmentation du Kd : ainsi, Gutierrez et Fuentes (1991) ont observé, pour des concentrations de cobalt de 1 mg.L-1 dans la solution du sol, des Kd respectifs de 151 et 318 L.kg-1 sec pour des sols différant essentiellement par leur teneur en argile, i.e. 16 et 31 %. La migration du cobalt dépend des caractéristiques du sol : en termes de pH, la mobilité diminue parallèlement à l'acidité ; dans les sols à faible capacité d'échange cationique (sablonneux acides, par exemple), le lessivage est modéré ; les sols ou horizons riches en matière organique fixeraient plus fortement et durablement le cobalt. La fraction remobilisable est dans ce cas d'environ 5 % par an avec une période (demi-vie) dans la couche humifère variant de 2 à 12 mois (Colle et al., 1991 ; 1996). La structure du sol en place est également un facteur important : des phénomènes particuliers de migration préférentielle et une valeur de Kd plus forte du cobalt lorsqu'il est mesuré en système batch, par rapport à une migration en lysimètres ont été mis en évidence par Albrecht et al. (2003). 1.3.2. Le cobalt et les végétaux Le cobalt est un oligo-élément qui jouerait un rôle vis-à-vis des bactéries symbiotiques fixatrices d'azote et dont l'excès pourrait provoquer une diminution de chlorophylles et de l'absorption du fer (Chatterjee et Chatterjee, 2000). L'absorption du cobalt par les plantes est conditionnée par la concentration du cobalt mobile du sol, de celui présent sur les sites 9 échangeables et par sa forme chimique. Par analogie avec le nickel, le transporteur dans les plantes semble être l'histidine (Bresson et al., 2009). À l'exception de quelques plantes hyper-accumulatrices, la plupart des plantes terrestres ne montrent pas de bioconcentration du cobalt. Ses teneurs moyennes dans les plantes vont de 0,1 à 115 mg.kg-1 sec. Des concentrations 10 à 20 fois plus élevées ont été observées dans des légumineuses sur des sols hydromorphes (Coughtrey et Thorne, 1983) ; sur des sols riches en cobalt, les concentrations des végétaux naturels peuvent atteindre de 100 à 1 000 mg.kg-1sec, notamment chez l'espèce arborée Nyssa sylvatica (Chatterjee et Chatterjee, 2003). Les facteurs de transfert racinaire moyens sont de l'ordre de 0,1 kg.kg-1 (poids secs). Ils varient entre 10-4 et 5. La variabilité pour un même type de végétal est en moyenne de l'ordre d'un facteur 10 et elle peut être beaucoup plus forte selon le sol et entre différents végétaux. L'absorption racinaire est réputée -- comme pour la plupart des cations -- augmenter faiblement avec la diminution du pH, ce qui apparaît dans les résultats de Gerzabek et al. (1994) ; à l'inverse, les résultats de Ban-nai et Muramatsu (2002) sur des radis cultivés sur 10 sols modérément acides (pH de 5,5 à 6,5) et ceux de Gerzabek et al. (1998) ne font pas apparaître de variation significative du transfert racinaire avec le pH du sol. Le fer, l'aluminium et le calcium influent sur l'absorption du cobalt par les végétaux par compétition sur les sites d'échange du sol (capacité d'échange). Dans une moindre mesure, la texture du sol et sa teneur en matière organique modifient l'intensité des transferts racinaires (Colle et al., 1996 ; AIEA, 2010). Le transfert aux dicotylédones est supérieur (en moyenne d'un facteur 15) au transfert aux monocotylédones. L'influence d'un apport de cobalt stable au sol a été mesurée lors des essais de Colle et al. (1991) en lysimètres de grande taille (16 bacs x 200 kg de sol brun calcaire). Les conclusions montrent qu'un apport de cobalt stable au sol entraîne une augmentation du facteur de transfert racinaire. Pour un amendement de 50 g de cobalt stable par kg de sol sec, le transfert du 60Co est approximativement multiplié par 8 pour les pailles de céréales et les gousses de haricots, par 4 pour les grains de céréales et par 2 pour les feuillages de légumes (salades, haricots, carottes). Un dépôt à la surface du sol augmente également le transfert comparativement à un apport incorporé de façon homogène dans la couche racinaire (Colle et al., 1983 ; 1996). L'étude de l'influence du mode d'apport montre que le dépôt global sur les organes aériens (aspersion sur les feuillages) est de loin le plus pénalisant, puisqu'il aboutit à une augmentation des facteurs de transfert (exprimés en Bq.kg-1 de végétal par Bq.m-2 de surface cultivée) d'un facteur 102 à 103 relativement à un apport racinaire de même activité surfacique équivalente (Colle et al., 1991). Ces résultats sont cohérents avec ceux de Sabbarese et al., (2002) qui ont comparé les activités massiques de tomates irriguées par aspersion et à la 10 raie : le ratio est de 6,5 environ pour les fruits, de 6,2 pour les tiges, de 5,3 pour les feuilles et de 2 pour les racines. Lors d'une contamination foliaire occasionnelle, les résultats, rares, sont en outre très variables selon le stade végétatif lors de la contamination. Lors d'expérimentations sur les céréales contaminées par aspersion à divers stades végétatifs, Coughtrey et Thorne (1983) indiquent, classiquement, que le pourcentage initial de radionucléide retenu par le végétal au moment de la contamination dépend de la biomasse à ce stade alors que le pourcentage retenu par le végétal à la récolte est lié au délai séparant la récolte de la contamination. La contamination du grain passe par un maximum lorsque la contamination a lieu pendant la période de maturation puis diminue, alors que celle de la paille augmente lorsque la date de contamination s'approche de celle de la récolte. La translocation vers le grain s'effectuerait ainsi durant une courte période. 1.3.3. Toxicité du cobalt dans les plantes Le cobalt à des concentrations élevées provoque une cytotoxicité et une phytotoxicité dans les plantes, qui sont similaires à celles du Cu, du Ni et du %00Zn. La cytotoxicité est l'inhibition de la mitose et l'endommagement des chromosomes, ainsi que la perturbation du réticulum endoplasmique des cellules de l'extrémité des racines (Rauser, 1981 ; Smith et Carson, 1981 ; Akeel et Jahan, 2020). La phytotoxicité varie en fonction des espèces végétales et de la concentration de Co dans les organes végétaux. Les légumineuses présentent généralement une chlorose ou une couleur blanc pâle sur les jeunes feuilles, et les tomates présentent soit une chlorose internervaire, soit une chlorose diffuse sur les jeunes feuilles (Akeel et Jahan, 2020). La toxicité du cobalt pour les plantes est rare dans les sols naturels, mais elle se produit lorsque les plantes poussent dans des sols contaminés par le Co. La contamination des sols par le Co provient principalement des activités d'extraction et de fusion, de l'élimination des boues d'épuration et de l'utilisation d'engrais chimiques (Hamilton, 1994). Comme indiqué ci-dessus, les plantes peuvent contrôler l'absorption, le transport et la distribution du Co. Cependant, lorsque le Co dans les sols contaminés devient hautement disponible, le Co peut obtenir un avantage compétitif sur le Fe, ce qui entraîne une plus grande absorption de Co que de Fe par l'IRT1. Avec des concentrations croissantes de Co à l'intérieur des cellules, FPN2 peut ne pas être en mesure de séquestrer efficacement le Co dans la vacuole, ce qui entraîne une plus grande quantité de Co à transporter des racines aux pousses. 11 1.4. Généralités sur la culture de blé Le blé est un terme qui désigne plusieurs céréales appartenant au genre triticum, ce sont des plantes annuelles cultivées dans de très nombreux pays. Le mot « blé » désignait également le « grain » (caryopse) produit par ces plantes. Le blé tendre (Triticum aestivum) et le blé dur (Triticum durum) sont les deux espèces les plus cultivées du genre Triticum dans le monde et le blé tendre fait partie des trois grandes céréales avec le maïs et le riz dont il est plus consommé par l'homme avec le riz et la troisième culture par l'importance de la récolte mondiale (Tavernier et Lizeaux, 1993). De tous les types de blés, deux ont une importance économique majeure : Le blé dur (Triticum durum) est surtout cultivé dans les régions sèches et chaudes au sud de l'Europe et en Afrique du Nord. Riche en gluten, il est employé pour les semoules et pâtes alimentaires (Clerget, 2011). Le blé tendre ou froment (T. aestivum), le plus cultivé, est produit dans les zones plus tempérées comme le nord de l'Europe et du continent américain pour la confection de la farine panifiable (Sabbagh, 2006). 1.4.1. Origine du blé Il y a 10 000 ans, à la fin de la dernière glaciation, des blés proches de ceux que nous cultivons aujourd'hui poussaient sur de vastes surfaces au Moyen-Orient, qui remonte au néolithique (environ 5000 ans avant J.C.) et peut-être même au mésolithique (7000 avant J.C), (Erroux, 1961). - Origine géographique L'origine géographique des blés est un des point les plus discutés ; à ce sujet plusieurs théorie et hypothèses ont été émises (Valdeyron, 1961). En effet selon Laumont et Erroux (1961), les recherches effectuées depuis fort longtemps sur le centre d'origine des blés ; basées sur des arguments archéologiques et phylogénétiques, permettant d'admettre que les trois groupes d'espèces du genre Triticum aurait trois centres d'origine distincts. Selon (Vavilov, Auriau, 1967 et Moule, 1980) ces groupes sont repartie comme suit : ? Groupes des Diploïdes : dont le centre d'origine est le foyer SYRIEN et le nord PALISTINIEN. ? Groupes des Tétraploïdes : ayant comme centre d'origine l'ABYSSINIE. 12 ? Groupes des Hexaploïdes : dont le centre d'origine est le foyer AFGHANO-INDIEN. Pour (Grignac ,1978), le moyen orient ou coexistent, les deux espèces parentales et où l'on a retrouvé de nombreuses forme de blé dur, serait le centre géographique. A partir de cette zone d'origine, l'espèce s'est différenciée dans trois centre : le bassin occidental, la méditerranée, le sud de l'ex URSS et le proche orient. L'Afrique du nord est considérée comme un centre secondaire de diversification de l'espèce (Bensemra, 1990). La diffusion des blés vers l'Afrique par la route la plus ancienne gagna l'Égypte vers 6 000 avant aujourd'hui et se poursuivit vers le Soudan et l'Éthiopie, au sud, et vers la Libye à l'est. D'autres voies d'introduction furent maritimes : à partir de la Grèce et de la Crète, certains blés rejoignirent également la Libye ; d'autres, en provenance du Sud de la péninsule italienne et de la Sicile, parvinrent aux côtes de la Tunisie, du Maroc et de l'Algérie (Bonjean, 2001). - Origine génétique Les espèces de blé tirent leur origine génétique de croisements naturels entre Triticum monococcum, Triticum urartu et des espèces sauvages apparentées appartenant à Aegilops (Aegilops speltoïdes) ; Triticum monococcum et Triticum urartu sont les premières formes de céréales cultivées, elles sont de constitution génomique 2n = 14. (Feldman et Sears, 1981). 1.4.2. Classification botanique et description morphologique - Classification botanique : Le blé est une plante herbacée, monocotylédone appartenant au groupe des céréales à paille. Il est une monocotylédone classée de la manière suivante : 13 Tableau 2. Classification systematique du blé selon (Van Den Abeelz, 1956)
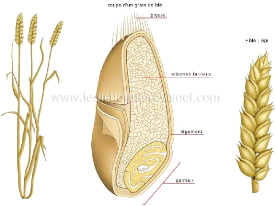
- Description morphologique : ? Le grain de blé : le grain est un caryopse, c'est un fruit sec indéhiscent, il est de couleur jaune ambrée à violacée selon l'espèce blé dur ou blé tendre et selon la variété. Il présente une partie plane (ventrale) et une partie dorsale légèrement bombée. La base élargie contient le germe est le sommet est garni de petits poils (la brosse), la partie ventrale est fondue par un sillon qui pénètre profondément dans le grain, cependant la partie dorsale présente une arête plus ou moins prononcée (Gond et al., 1986 in Oudjani, 2009). Chez le blé dur comme la plupart des graminées possèdent un fruit sec (caryopse), celle-ci distingué par un grain étroit allongé à sillon profond à brosse peu développé et à texture souvent vitreuse (Bachir Bey Ilhem et al 2015). Elle est entourée d'une matière végétale qui la protège des influences extérieures. Elle est constituée d'un germe ; la partie essentielle du fruit permettant la reproduction de la plante: il se développe et devient à son tour une jeune plante et d'une amande avec 65 à 70% d'amidon ainsi qu'une substance protéique (le gluten) dispersée parmi les grains d'amidon (Bebba 2011). 14
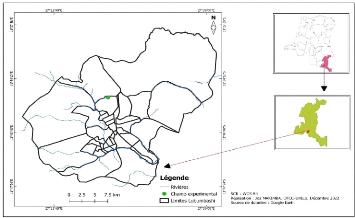
Figure 2. Anatomie du grain de blé ? La tige : La tige commence à prendre son caractère au début de la montaison, c'est-à-dire prend sa vigueur et porte 7 à 8 feuilles, elle présente des bourgeons auxiliaires que servent à l'origine des talles, elle s'allonge considérablement à la montaison. (Alismail et al., 2017). ? Les feuilles : les feuilles sont à nervures parallèles. Le limbe possède souvent à la base deux prolongements aigus embrassant plus ou moins complètement la tige : les oreillettes ou stipules ; à la soudure du limbe et de la gaine peut se trouver une petite membrane non vasculaire entourant en partie le chaume (Belaid, 1986). La feuille terminale a un rôle primordial dans la reproduction (Soltner, 1988). ? Les racines du blé sont fibreuses : à la germination la radicule ou racine primaire, et un entre-noeud sub-coronal émergent du grain. Le système racinaire secondaire peut être assez développé, s'enfonçant à des profondeurs atteignant jusqu'à deux mètres. Il apporte les éléments nutritifs à la plante (Soltner, 1988). Les racines de blé sont de type fasciculé peu développé dont on peut distinguer des racines primaires qui assurent la croissance de la plantule jusqu'au tallage tandis que les racines secondaires ou adventices sont émises à partir du plateau de tallage. Celles produites par la plantule durant la levée sont des racines séminales tandis que ces adventives se forment plus tard à partir des noeuds à la base de la plante et constituent le système racinaire permanent (Maamri, 2011). La profondeur des racines est variable selon le travail du sol ainsi que l'humidité du sol. Pour Bachir Bey et al., (2015), 50% du poids total des racines se trouvé entre 0 - 25 cm, 10% entre 20 - 50 cm mais elles peuvent atteindre 1m à 1.2m 15 dans un sol bien profond. Cependant pour Fritas, (2012), 55 % du poids total des racines se trouve entre 0 et 25 cm de profondeur. ? Les fleurs : sont regroupées en une inflorescence composée d'unités morphologiques de base : les épillets. Chaque épillet compte deux glumes (bractées) renfermant de deux à cinq fleurs distiques sur une rachéole (Soltner, 1988). Elles sont groupées en inflorescence ou épillets qui s'attachent à l'axe ou rachis de l'épi portant de 15 à 25 épillets par épis (Benderradji, 2013) et comportant de 3 à 5 fleurs, tandis que chaque fleur est enveloppée de deux glumelles l'une à l'intérieur et l'autre à l'extérieur dont chacune compte trois étamines à anthères biloculaires, ainsi qu'un pistil à deux styles à stigmates plumeux qui peut produire un fruit à une seule graine, soit le caryopse (Maamri, 2011). Au cours de la fécondation, les anthères sortent des fleurs tandis que le grain est à la fois le fruit et la graine (Bebba 2011). La fécondation de la fleur a lieu à l'intérieur des glumelles, avant la sortie des étamines à l'extérieur (Benderradji ; 2013). 1.4.3. Cycle biologique du blé 1. La période végétative Elle se caractérise par un développement strictement herbacé et s'étend du semis jusqu'à la fin de tallage (Bada., 2007). - La germination-levée Cette phase comprend une période de l`imbibition de la graine, libération des enzymes et dégradation des réserves assimilables par la graine, ensuite c'est la phase de croissance caractérisée par l`allongement de la radicule (Hennouni, 2012). C'est un passage de la semence de l'état de vie lente à l'état de vie active (Benchikh , 2015). Le grain de blé absorbe au moins 30% de son poids en eau (Fritas, 2012). Juste à la levée, les premières feuilles déclenchent la photosynthèse. On parlera de levée lorsque 50 % des plantes seront sorties du sol (Ait Slimane AitKaki, 2008). Les téguments se déchirent, la racine principale est couverte d'une enveloppe appelée coleorhize apparaît, suivie par la sortie de la première feuille, couverte d'une enveloppe appelée coléoptile (Fritas., 2012). Ces organes jouent un rôle protecteur et mécanique pour percer le sol (Ait Slimane AitKaki, 2008). Au cours de la germination la coléorhize s'épaissit en une masse blanche et brise le tégument de la graine au niveau du germe, c'est le début de l'émission des racines primaires, garnis de poils absorbants, tandis que le coléoptile gainant la première vraie feuille, s'allonge vers la 16 surface, où il laisse percer la première feuille, c'est la levée alors que la deuxième et la troisième feuille suivent bien après (Laala, 2011) et une tige sur le maitre brin à l'aisselle de la feuille la plus âgée se lance à la surface du sol (Belagrouz, 2013), puis apparaissent d'autres racines et feuilles. La durée de cette phase varie avec la température de 8 à 15 jours (Fritas, 2012). La date de levée est définie par l'apparition de la première feuille qui traverse la coléoptile, cependant la germination de la graine dépend de trois facteurs importants ; l'eau, l'aération et la température (l'optimum de la germination de 15/25°C) (Bachir Bey, et al 2015). Le blé germe dès que la température dépasse le 0 °C (Bebba, 2011). Le rythme d'émission des feuilles est réglé par des facteurs externes comme la durée du jour et la température tandis que la somme de température séparant l'apparition de deux feuilles successives est estimée à 100°C et varie entre 80 °C pour le semis tardif et à 110 °C pour un semis précoce (Bebba 2011). Les plus grosses graines lèvent les premières et donnent des plantules plus vigoureuses cependant, la composition des réserves (teneur en protéines) agit favorablement sur la vitesse de la germination-levée (Nadjem, 2011). - Tallage Cette phase s'amorce à partir de la quatrième feuille. La formation de la première talle se fait au stade 3 feuilles. La première talle primaire (maitre-brin) apparaît à l'aisselle de la première feuille du blé. La 2ème et la 3ème talle apparaissent à l'aisselle de la 2ème et la 3ème feuille (Hamadache, 2013 et Salmi, 2015). Le fin tallage est celle de la fin de la période végétative, elle marque le début de la phase reproductive, conditionnée par la photopériode et la vernalisation qui autorisent l'élongation des entrenoeuds (Gate, 1995 et Salmi, 2015). Le début du tallage est caractérisé par : ? la formation de nouvelles racines (la plante possède de 5 à 6 racines primaire) (Bachir Bey et al., 2015). ? stade de formation du plateau de tallage : c'est le phénomène de "pré-tallage" dans lequel le deuxième entre noeud qui porte le bourgeon terminal s'allongé à l'intérieur de la coléoptile, il cesse de remonter à 2 cm sous la surface (Laala, 2011), quelle que soit la profondeur du semis, à ce niveau on assiste à l'apparition d'un renflement : c'est le futur plateau de tallage (Benchikh, (2015) il se forme presque au niveau de la surface du sol ( Bachir Bey et al., 2015). ? stade d'émission des talles : Des valeurs de la variance fortement significatives et positives pour le nombre d'épillets par épi ont été retrouvées chez tous les croisements 17 effectués par Emir (2010) et qui exhibent aussi un hétérobeltiosis significatif et positif. Bachir Bey et al., (2015) à l'aisselle des premières feuilles du blé des bourgeons axillaires entre, alors en activité pour donner de nouvelles pousses: les talles. Bencheikh, (2015), la première talle se forme à la base de la première feuille et la deuxième talle à la base de la deuxième feuille. Ouanzar (2012), les bourgeons axillaires à l'aisselle des feuilles des talles donnent naissance à l'émission de talles secondaires. Il apparaît à partir de la base du plateau de tallage, des racines secondaires ou adventives, qui seront à l'origine de l'accroissement du nombre d'épis. Le stade fin tallage c'est-à-dire au stade "épi à 1 cm du plateau de tallage, est caractérisé par une croissance active des talles. Le plant de blé a besoin, durant cette phase, d'un important apport d'engrais azotés (Bebba 2011). L'importance du tallage dépendra de la variété de, la densité de semis, la densité d'adventices et de la nutrition azotée cependant, le tallage marque la fin de la période végétative et le début de la phase reproductive, qui est conditionnée par la photopériode et la vernalisation qui autorisent l'élongation des entre-noeuds (Ait Slimane AitKaki, 2008). 2. Période de reproduction - Phase de la montaison Au cours de cette phase pour Fritas (2012), les talles montantes entrent en compétitions pour les facteurs du milieu avec les talles herbacées qui de ce fait n'arrivent pas à monter en épis à leur tour. Ces dernières régressent et meurent (Laala, 2011). Ce phénomène se manifeste chez un certain nombre de talles herbacées qui commence (jeunes talles) par une diminution de la croissance puis par un arrêt de celle-ci alors que, d'autres se couronnent par des épis (Laala, 2011). Pendant cette phase de croissance active, les besoins en éléments nutritifs notamment en azote sont accrus (Ait Slimane AitKaki, 2008). Non seulement deviennent très importants mais aussi déterminent le nombre d'épis, le nombre de grain par épi et le poids maximal du grain (Benchikh 2015). La montaison s'achève à la fin de la sortie de la dernière feuille et le début du gonflement qui engaine les épis dans la gaine (Nadjem, 2011). - Phase de l'épiaison et de fécondation Elle est marquée par la méiose pollinique, l'éclatement de la gaine avec l'émergence de l'épi. C'est au cours de cette phase que s'achève la formation des organes floraux et s'effectue la fécondation (Soltner, 2005). 18 La vitesse de croissance de la plante est maximale. Cette phase correspond à l'élaboration d'une grande quantité de la matière sèche, cette phase dépend étroitement de la nutrition minérale et de la transpiration qui influence le nombre final de grains par épi (Masale, 1980 ; Soltner, 2005). - La maturation du grain Au cours de cette phase, l'embryon se développe et l'album se charge de substances de réserve. On observe une augmentation du volume du poids des graines (stade laiteux). Ensuite, le poids frais des grains continue à augmenter alors que celui des tiges et des feuilles diminue (stade pâteux). Puis les grains deviennent durs et leur couleur devient jaunâtre (Boufnar-Zaghoune et Zaghouane, 2006). I.1.5. les exigences climatiquesPour être rentable, la production des céréales nécessite une connaissance accrue du développement de la culture. Selon Djermoun, (2009) les facteurs climatiques ont une action prépondérante sur les différentes périodes de la vie du blé. Parmi ces facteurs on citera principalement : - Température La germination commence dès que la température dépasse 0°C, avec une température optimale de croissance située entre 15 à 22° C. Les exigences globales en température sont assez importantes et varient entre 189o et 24 °C selon les variétés. De même la température agit sur la vitesse de croissance, elle ne modifie pas les potentialités génétiques de croissance, c'est la somme de température qui agit dans l'expression de ces potentialités. Chaque stade de développement du blé nécessite des températures particulières (Ondo., 2014). - L'humidité Les besoins en eau de la culture du blé varient entre 450 et 650 mm selon le climat et la longueur du cycle végétatif (Baldy, 1974). - La photopériode Le rayonnement solaire et la durée du jour consolide l'effet positif de la température sur le rendement quand elle n'est pas très élevée et accentue son effet négatif dans le cas contraire. La photopériode affecte aussi la durée de chacune des périodes de développement citées. Les variétés du blé diffèrent quant à leur sensibilité à la photopériode (Kalarasse., 2018). 19 - L'eau L'eau est un facteur limitant de la croissance du blé. Ce dernier exige l'humidité permanente durant tout le cycle de développement. Pendant les différentes phases de son cycle, les besoins sont plus élevés au vu des conditions climatiques défavorables (Bennai et Benabbas, 2007). - La lumière La lumière est le facteur qui agit directement sur le bon fonctionnement de la photosynthèse et le comportement du b lé. Un bon tallage est garanti si le blé est placé dans les conditions optimales d'éclairement (Felliet, 2000). 1.4.4. Importance de la culture Historiquement, le blé est l'une des premières céréales cultivées et consommées dans le monde avec une production annuelle d'environ 600 millions de tonne (Ricroch et al., 2011). Les échanges qui se multiplient entre les régions du monde font de cette céréale l'un des principaux acteurs de l'économie mondiale (Laberche, 2004). La production du blé est surtout localisée dans le bassin méditerranéen d'une part (Europe du Sud, Moyen orient, Afrique du Nord), et en Amérique du Nord d'autre part (Canada central et Nord des USA), où est produit le quart du blé dur mondial (Clerget, 2011). Depuis la nuit des temps les céréales nourrissent l'humanité, produire mieux et plus a été nécessaire pour répondre aux besoins d'une population grandissante. Nos ancêtres ont dû apprendre à améliorer progressivement les techniques de production, sélectionner des semences avec un meilleur potentiel génétique. Depuis longtemps, les céréales, notamment le blé est devenu un produit de première nécessité à l'échelle mondiale. Son importance dépasse le rôle traditionnel considéré comme aliment (Ammar, 2015). L'amidon du blé tendre est également utilisé depuis plusieurs années comme matière première pour la fabrication de biocarburants (Clément, 2010). En 2018, la production mondiale de blé a atteint 762 millions de tonnes contre 732 millions de tonnes pour 2019 (CIC, 2019). Au cours la compagne agricole 2018-2019, les principaux producteurs de blé mondiaux sont, par ordre décroissant, sont l'Union Européenne (137.600 milliers de tonnes), la Chine (131.430 milliers de tonnes), l'Inde (99.700 milliers de tonnes), la Russie (71.600 milliers de tonnes), les Etats Unies (51.287 milliers de tonnes) et le Canada (31.800 milliers de tonnes) (Statista, 2019). Le marché mondial du blé est segmenté en différents groupes de pays qui ont diverses capacités de production et de consommation de blé, ce qui rend ce marché plus propice à la volatilité des 20 prix. Seulement 20% de la production mondiale du blé est échangée et il s'agit d'un marché de surplus et d'excédents. Cependant ce commerce mondial du blé a été multiplié par trois entre 1961 et 2005 et le blé conforte ainsi sa place de céréale « la plus échangée » dans le monde. (CIC, 2019). 21 Chapitre 2. Milieu, matériels et méthodes 2.1. Milieu d'étude 2.1.1. Situation géographique Le présent travail a été réalisé dans la région de Lubumbashi, située au Sud-est dans la province du Haut-Katanga en République démocratique du Congo ; à une altitude d'environ 1250 m et elle est située entre 11° 20' et 12° de latitude Sud et 27° 10' et 27° 40' de longitude Est.
Figure 3. Localisation du milieu d'étude 2.1.2. Climat La ville de Lubumbashi appartient au type du climat Cw6 d'après la classification de Koppen. Ce climat est caractérisé par une alternance de deux saisons durant 6 mois chacune avec une période de transition au mois d'Avril (la saison des pluies et la saison sèche). Les précipitations annuelles varient entre de 1270 mm et de 1770 mm (FAO, 2005). Le climat de la ville de Lubumbashi est donc tropical humide, caractérisé par une alternation des deux saisons : (1) la saison sèche qui dure 6 mois et 7 mois ; elle débute, en moyenne, le 20 avril et prend fin vers le 20 octobre ; (2) la saison des pluies qui dure 6 mois, du mois d'Octobre à Avril (soit 230 à 240 jours de pluie ; Kasongo, 2009 ; Mukalay, 2016). Durant la saison des pluies, il tombe une moyenne de 1200 mm, ce qui signifie qu'il ne pleut presque pas durant la saison sèche. 22 La température moyenne annuelle varie autour de 20°C. Les mois les plus chauds sont ceux d'octobre et novembre avec une moyenne maximum journalière de 32°C et une température moyenne mensuelle de 23°C. Par contre, le mois de juillet est le mois le plus froid avec la moyenne minimum journalière de 8°C, la température moyenne mensuelle étant de 17°C (FAO, 2005 ; Mpundu, 2010). Le taux d'humidité moyenne annuelle est de 68 % avec les niveaux d'humidité mensuelle dont les moyennes varient d'un minimum de 47% en saison sèche (septembre) et un maximum de 86% durant la saison des pluies (février) (Mpundu, 2010). 2.1.3. Sol Le Haut-Katanga, étant une province minière, est caractérisée par les sols polygéniques, qui proviennent principalement du socle précambrien de Lubumbashi synclinal dominé en grande partie des schistes de la série de Kundelungu (Kasongo, 2009). Les sols du Haut-Katanga font partie des sols latéritiques désaturés jaune-rouge. Ils sont caractérisés par une abondance des matériaux argileux (Kaolinite) avec une faible capacité d'échange cationique (CEC), du Fer, des sesquioxydes d'Aluminium et un pHeau flottant à 5,2 (Kasongo et al., 2013). Les sols du Haut-Katanga sont très diversifiés par leur nature. La carte des sols élaborée par l 'ISRIC (Van Engelen et al., 2006 in Kasongo 2009) dans le cadre de la base des données SOTER montre, la dominance de groupes des sols de référence suivants : Ferralsols, Vertisols, Cambisol et Fluvisols. Les Ferralsols orthiques, Vertisols, Combisols et Gleysols sont les principaux groupes qui constituent les différentes classes des sols dans le Haut-Katanga, ils font partie des sols tropicaux récents et terres noires tropicales de la classification de l'INEAC (Kasongo, 2009). Le relief est caractérisé par les chaines des montagnes de Mitwaba, Kundelungu et d'Upemba. 2.1.4. Hydrographie Ville située sur plusieurs confluents. Le principal, plus proche du centre historique et définissant la position du plateau sur lequel est posée la ville, est celui de la Kafubu avec la Lubumbashi, son affluent, au sud du noyau urbain originel. Les autres confluents dans l'agglomération ont lieu entre la Lubumbashi et la Karavia au centre-ouest de la ville, la Kafubu et la Ruashi à l'est, et entre la Kafubu et la Naviundu, au sudouest. Des dembos (savannes saisonnièrement inondées) sont formés dans certains fonds de vallée des petits affluents. Un lac de retenue a été aménagé sur la Lubumbashi, dans la partie nord de l'agglomération (Vranken, 2010). 23 2.1.5. Végétation Les principales unités de végétation dans le Haut-Katanga sont : les forêts denses sèches, les forêts denses édaphiques, les forêts claires, les savanes et les milieux aquatiques (Muganguzi, 2016). Le Haut-Katanga et ses environs sont caractérisés par des savanes arborées anthropiques, ce qui entraîne la dégradation des forêts de Miombo en savane et arbustive et finalement en savane herbeuse (Malaisse, 1997). La forêt claire du type Miombo s'impose comme l'écosystème le plus dominant, il a fait l'objet des nombreuses études, tant botaniques, phytosociologiques, zoologiques qu'écologiques (Meerts et Hasson, 2016 ; Malaisse, 2018). Les forêts dans le Haut-Katanga se distinguent en quatre types principaux ci-après : la forêt claire (Miombo), la forêt dense tropicale (Mushitu), et la forêt sèche sempervirente (Muhulu) (Malaisse et al., 1972), et les forêts denses marécageuses des hauts plateaux qui sont méconnues (Malaisse, 2018). La forêt claire à Brachystegia appelée « Miombo » est le biome dominant caractérisé par de termitières occupant aussi une grande surface terrière, elle couvre plus de 80% du territoire et constitue une ressource économique très importante pour la province (Mujinya et al., 2014). 2.2. Matériels 2.2.1. Matériels végétal > La variété du blé choisie appelée « 59/26 » est une variété qui provient du Mexique, elle a un cycle végétatif de 120 jours, elle peut atteindre une hauteur de 120cm et donne un rendement de 1 à 2t/ha. 2.2.2. Matériels non biologiques > Une latte > Des pots avec sous-plats > Un stylo 2.2.1. Matériels chimiques Pour notre étude nous avons utilisé les réactifs ci-après que nous avons contaminés au sol. > Le sulfate du cuivre pentahydraté : CuSO4 5H2O > Le sulfate de cobalt heptahydraté : CoSO4 7H2O Figure 4. Schéma du dispositif expérimental adopté 24 2.3. Méthodes Deux essaies séparés sont adoptés, l'un pour le Cu et l'autre pour le Co. les substrats reçoivent 4 traitements en utilisant le sulfates de cuivre pentahydraté (CuSO4 5H2O) et le sulfates de cobalt heptahydraté (CoSO4.7H2O), elles sont mélangées directement au sol dans les pots. Les doses initiales étaient respectivement de T0 : 0 ppm ; T1 : 50 ppm ; T2 : 100 ppm ; T3 : 200 ppm (mg.kg-1), lorsque les grains ont été mises en terre, il n'y avait aucun grain qui a germé sauf dans le traitement témoin, sur ce nous avons dilué la concentration de chaque traitement 3 fois, et nous avons eu des concentrations diluées suivantes : T0 : 0 ppm ; T1 : 16 ppm ; T2 : 33 ppm ; T3 : 66 ppm. Tableau 3. Quantité de sulfates de cuivre
pentahydraté et de sulfates de cobalt
Le dispositif expérimental adopté est en Blocs aléatoires complets et comporte quatre traitements (4), avec trois (3) répétitions.
BLOC 1 BLOC 2 BLOC 3 T0R1 T1R1 T2R1 T3R1 T3R2 T2R2 T1R2 T2R3 T3R3 T0R3 T1R3 T0R2 T0 : 0 ppm ; T1 : 16 ppm ; T2 : 33 ppm ; T3 : 66 ppm. > Le nombre de feuilles par plantes : le nombre des thalles a été prélevé au 20eme et au 40eme jour. 25
Figure 5. Dispositif expérimentale adopté pour le CuSO4 5H2O et pour le CoSO4 7H2O séparément 2.3.1. Conduite de l'essai + Prélèvement du sol expérimental Le sol expérimental a été prélevé dans la foret da Miombo, et 4 échantillons composites ont été prélevé sur une superficie de 1ha, et les échantillons ont été homogénéiser et après conditionné dans les pots. + Le semis Nous avons effectué un semis direct dans les pots, et les pots avaient une capacité de contenir 1kg de sol, à raison de 4 grains par pot, disposé systématiquement. + L'arrosage En ce qui concerne l'arrosage, elle se faisait dans les sous-plats pour éviter le tassement du sol à la surface du sol. 2.3.2. Paramètres mesurés ? Paramètres morphologiques > Le taux de survie : le taux de survie en (%) a été prélevé au 15eme jour ainsi que au 30eme jour. > La hauteur des plantes : la hauteur des plantes en (Cm) a été mesure au 20eme et au 40eme jour. 26 ? La longueur des racines : après la récolte des plantes nous avons mesuré la longueur de racines en (Cm). 2.3.2. Analyses statistiques Les analyses de la variance (ANOVA) pour les paramètres végétatifs notamment (le taux de germination, le taux de survie, la hauteur de plantes, le nombre de feuilles, la longueur de racine) du blé ont été réalisées à l'aide du logiciel R. Le test de Tukey a été utilisé en vue de comparer les différentes moyennes à l'aide du logiciel MINITAB, nous avons utilisé Excel 2019 pour la réalisation des graphiques. 27 Chapitre 3. Interprétation des résultats 3.1. Résultats obtenus avec le traitement de Cu 3.1.1. Taux de germination (%) En ce qui concerne le taux de germination, l'analyse de la variance montre qu'il y a des différences significatives entre les différents traitements (P < 0,05). En effet, le taux de germination des grains de blé tendre tend à diminuer de manière inversement proportionnelle aux concentrations croissantes du Cu comme la figure (8) l'illustre. MOYENNE A 120
Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements Figure 6. Taux de germination à 10 jours en fonction des traitements Figure 8. Taux de survie à 40 jours en fonction des traitements 28 3.1.2. Taux de survie (%) L'analyse statistique révèle un effet significatif (P < 0,05) sur le taux de survie des plantes de blé tendre à 15 jours, la moyenne la plus élevée a été observée au traitement témoin avec une valeur de 100% et la moyenne les plus faibles ont été observées avec tous les traitements ayant reçu du Cu (figure 9). MOYENNE A 120 Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements
T.S 15 jours 100 40 80 60 20 0 B B B Figure 7. Taux de survie à 15 jours en fonction des traitements MOYENNE 120 Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements
T.S 40 jours 100 40 80 60 20 0 B B B A Egalement l'analyse statistique révèle une différence significative entre les différents traitements, en ce qui concerne le taux de survie à 40. La moyenne la plus élevée a été observé au traitement témoin comme la figure (10) l'illustre. Figure 10. Hauteur des plantes à 40 jours en fonction des traitements 29 3.1.3. Hauteur de plantes (Cm) La figure (11) met en évidence l'effet des différentes concentrations du Cu sur la hauteur moyenne des plantes de blé tendre après 20 jours de traitement. Une diminution nette de la hauteur des blés cultivés sur des traitements au Cu comme le montre la figure 11.
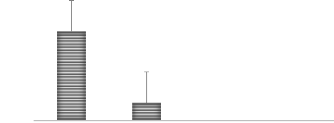
A 30 25 H.P 20 jours 20 15 10 5 0 B B B Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements MOYENNE Figure 9. Hauteur des plantes à 20 jours en fonction des traitements H.P 40 jours
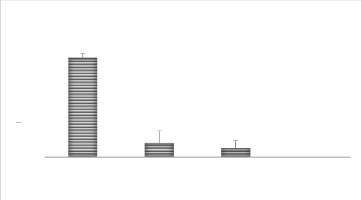
Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements MOYENNE 35 30 A Pour la hauteur de plantes à 40 jours, l'analyse statistique montre qu'il y a une différence significative entre les différents traitements témoin d'une part et tous les traitements avec le Cu.(P < 0,05), la moyenne la plus élevée étant observée dans le traitement témoin comme l'illustre la figure 12. Figure 12. Nombre des feuilles à 40 jours en fonction des traitements 30 3.1.4. Nombre de feuilles Pour le nombre de feuilles à 20 jours, l'analyse statistique montre qu'il y a plus de feuilles sur le blé du traitement témoin qu'il y en a sur les traitements au Cu. Une différence significative entre les deux tendances (P < 0,05), une fois de plus les traitements au Cu affiche la valeur la plus faible que celle du témoin (figure 13).
Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements MOYENNE 7 6 A Figure 11. Nombre des feuilles à 20 jours en fonction des traitements N.F 40 jours MOYENNE 7 A
Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements Au quarantième jour l'analyse statistique dégage une différence significative entre les traitements témoin et les traitements au Cu. Le nombre de feuilles du traitement témoin s'avère plus élevé que celles des traitements qui ont reçu le Cu. (Figure 14). 31 3.1.5. Longueur des racines (Cm) La figure (15) met en évidence l'effet des différentes concentrations du Cu sur la longueur moyenne des racines de blé tendre après 40 jours de traitement. Nous constatons une diminution de la longueur moyenne des racines inversement aux ayant reçu le Cu. Cette diminution est significative (P < 0,05) chez les plantés traitées avec le Cu. L.R 40 jours
Temoin 16 ppm de Cu 33 ppm de Cu 66 ppm de Cu Traitements MOYENNE 16 14 12 A Figure 13. Longueur des racines à 40 jours en fonction des traitements 32 3.2. Résultats obtenus avec le traitement Co 3.2.1. Taux de germination (%) En ce qui concerne le taux de germination, l'analyse de la variance montre qu'il y a des différences significatives entre les traitements témoin, la dose 1 et 2 d'une part et le traitement T3 (P < 0,05). En effet, le taux de germination des grains de blé diminue significativement dans la dose la plus élevée comme le montre la figure (16). MOYENNE A 120 Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements
T.G 10 jours 100 40 80 60 20 0 A A B Figure 14. Taux de germination à 10 jours en fonction des traitements Figure 16. Taux de survie à 40 jours en fonction des traitements 33 3.2.2. Taux de survie (%) L'analyse statistique révèle un effet significatif (P < 0,05) sur le taux de survie des plantes de blé tendre à 15 jours, la moyenne la plus élevée a été observée au traitement témoin tandis que les valeurs les plus faibles s'observent dans le traitement avec 33 ppm de Co et avec 66 ppm de Co et la valeur la plus élevée revient au traitement témoin. Le traitement avec 16 ppm étant intermédiaire entre ces deux tendances comme l'exprime la figure 17. MOYENNE A 120 Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements
T.s 15 jours 100 40 80 60 20 0 AB B B Figure 15. Taux de survie à 15 jours en fonction des traitements MOYENNE 120 100 A
Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements L'analyse de la variance montre une différence significative entre les traitements en ce qui concerne le taux de survie à 40 jours, et la moyenne la plus élevée a été observée au traitement témoin comme l'illustre la figure (18). 34 3.2.3. Hauteur de plantes (Cm) La figure (19) met en évidence deux tendances différentes concentrations du Co sur la longueur moyenne des racines de blé. La valeur la plus élevé s'observe dans les traitements témoin et les plus faibles dans les doses ayant reçu le Co. MOYENNE 35 30 Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements
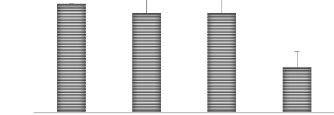
H.P 20 jours 25 20 15 10 5 0 B B B A Figure 17. Hauteur des plantes à 20 jours en fonction des traitements
Figure 18. Variation des hauteurs en fonction des traitements 35 Pour la hauteur de plantes à 40 jours, l'analyse statistique montre qu'il y a une différence significative entre les différents traitements (P < 0,05), la moyenne la plus élevée observée au traitement témoin et les moyennes les plus ont été observées avec les traitements (33 et 66 ppm). H.P 40 jours
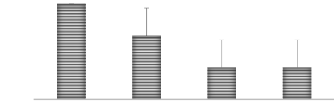
Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements MOYENNE 45 40 35 A Figure 19. Hauteur des plantes à 40 jours en fonction des traitements Figure 21. Nombre des feuilles à 40 jours en fonction des traitements 36 3.2.4. Nombre de feuilles Pour le nombre de feuilles à 20 jours, l'analyse statistique montre qu'il y a une différence significative entre les différents traitements (P < 0,05), la moyenne la plus élevée observée au traitement témoin et les moyennes les plus faibles ont été observées avec les traitements (33 et 66ppm de Co).
Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements MOYENNE 7 6 A Figure 20. Nombre des feuilles à 20 jours en fonction des traitements N.F 40 jours
Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements MOYENNE 14 12 A L'analyse statistique dégage une différence significative entre les traitements pour le nombre de feuilles à 40 Jours, et la moyenne la plus élevée a été observée au traitement témoin comme la figure (23) l'illustre. 37 3.2.5. Longueur des racines à 40 jours (Cm) La figure (23) met en évidence l'effet des différentes concentrations du Co sur la longueur moyenne des racines de blé tendre après 40 jours de traitement. Les résultats montrent une diminution de la longueur moyenne des racines inversement aux concentrations croissantes du Co par rapport aux témoins. Cette diminution est significative (P < 0,05) chez les plantés traitées avec par la concentration de 16, 33, et 66 ppm de Cu. MOYENNE 14 Temoin 16 ppm de Co 33 ppm de Co 66 ppm de Co Traitements
L.R 40 jours 12 10 4 8 6 0 2 B B B A Figure 22. Longueur des racines à 40 en fonction des traitements 38 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||